

Peripheral Nerve Regeneration Is Independent From Schwann Cell p75NTR Expression
- PMID: 31191256
- PMCID: PMC6548843
- DOI: 10.3389/fncel.2019.00235
Peripheral Nerve Regeneration Is Independent From Schwann Cell p75NTR Expression
Abstract
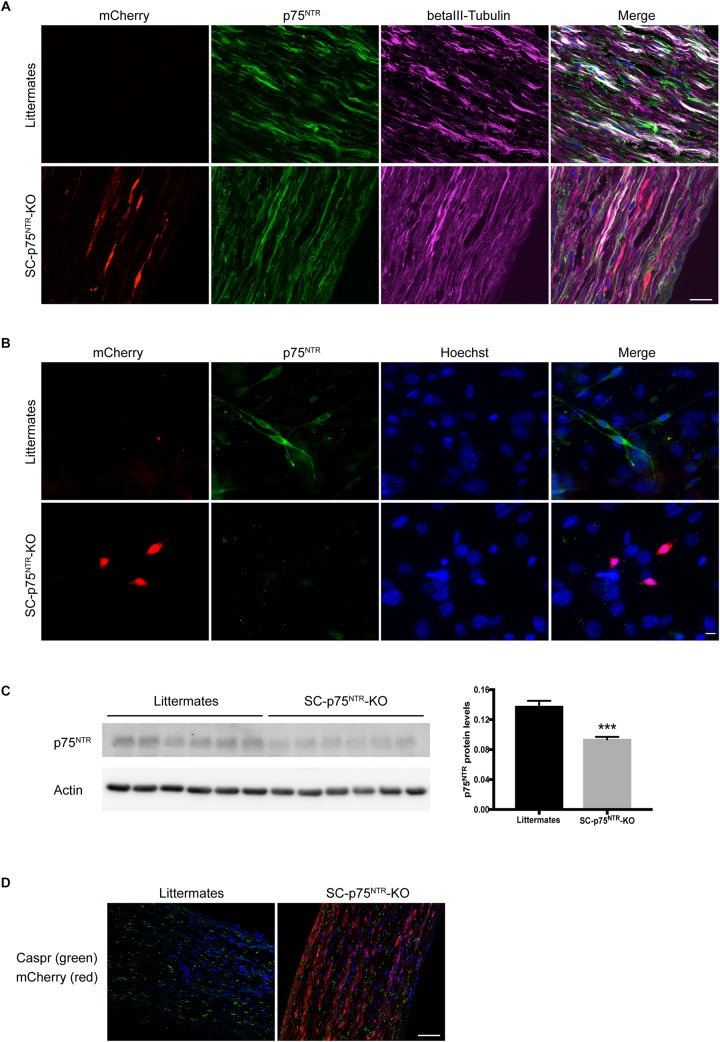
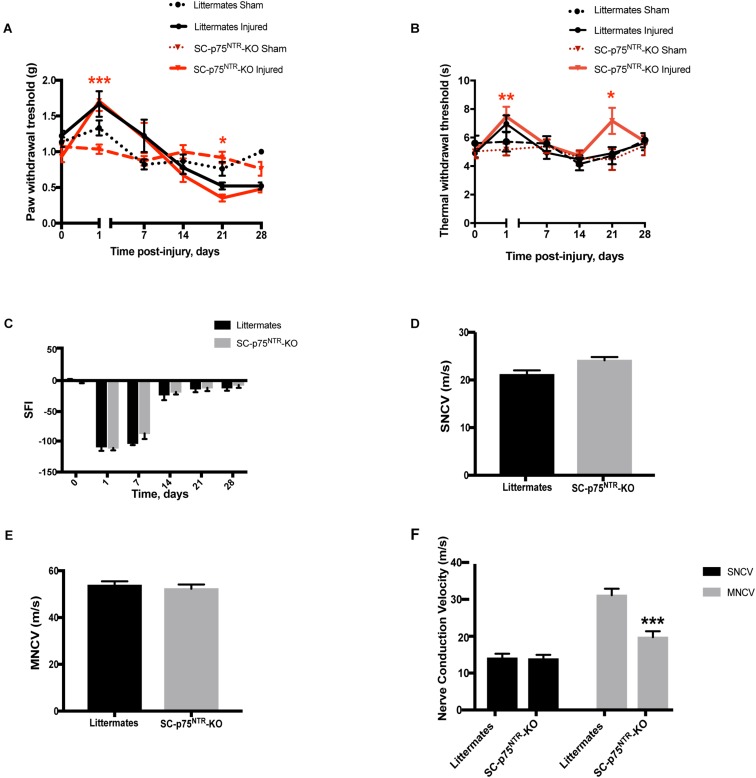
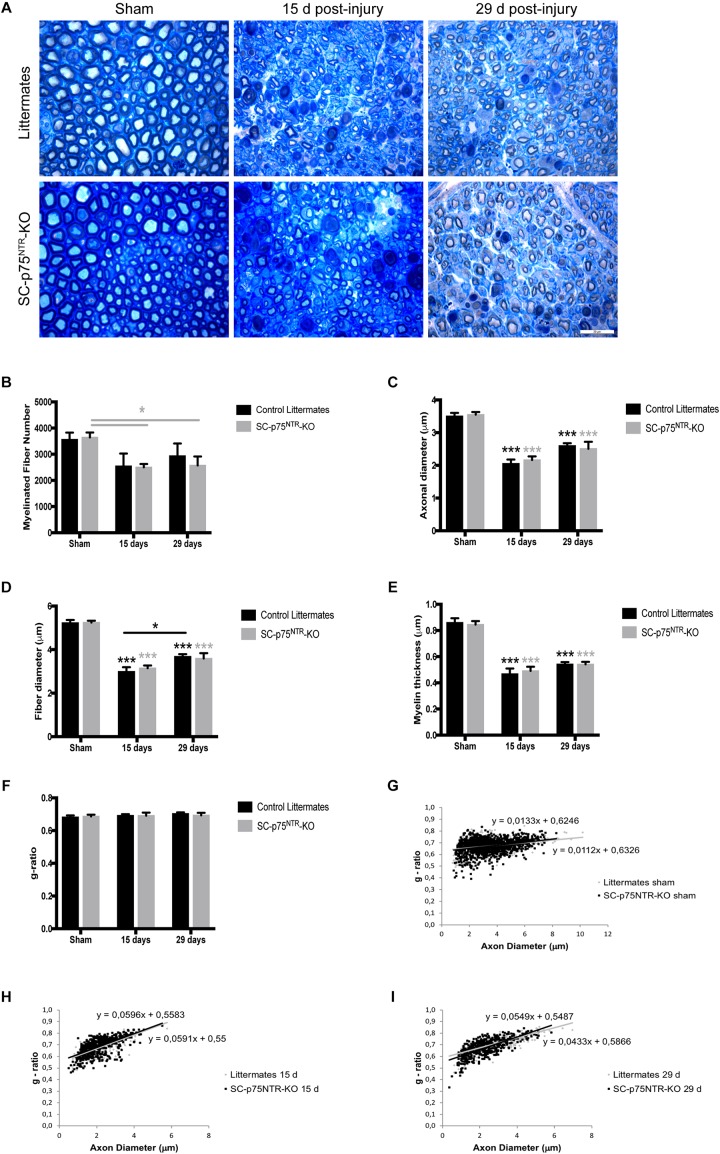
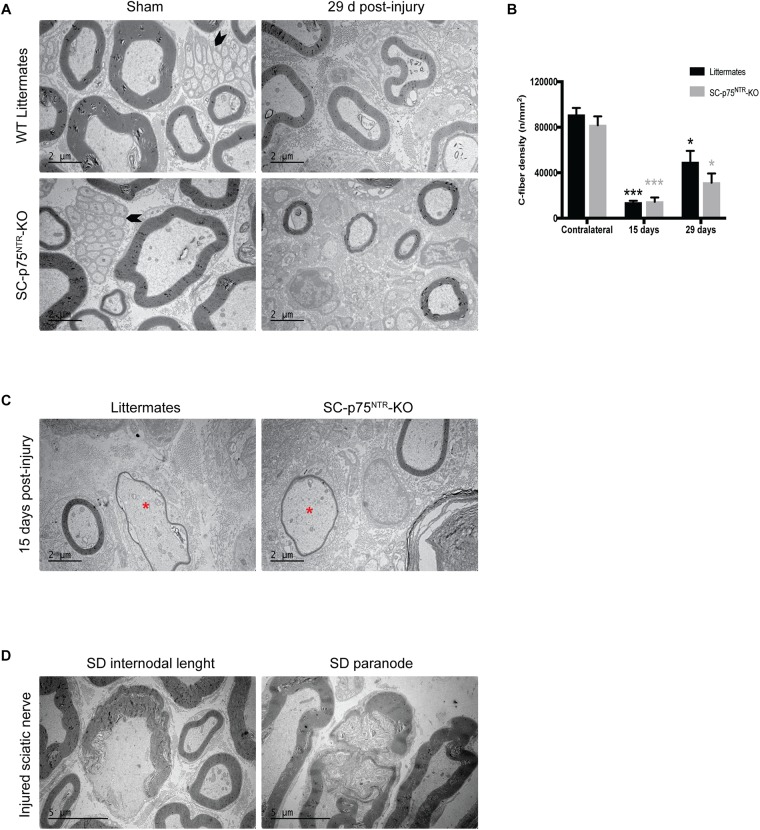
Schwann cell reprogramming and differentiation are crucial prerequisites for neuronal regeneration and re-myelination to occur following injury to peripheral nerves. The neurotrophin receptor p75NTR has been identified as a positive modulator for Schwann cell myelination during development and implicated in promoting nerve regeneration after injury. However, most studies base this conclusion on results obtained from complete p75NTR knockout mouse models and cannot dissect the specific role of p75NTR expressed by Schwann cells. In this present study, a conditional knockout model selectively deleting p75NTR expression in Schwann cells was generated, where p75NTR expression is replaced with that of an mCherry reporter. Silencing of Schwann cell p75NTR expression was confirmed in the sciatic nerve in vivo and in vitro, without altering axonal expression of p75NTR. No difference in sciatic nerve myelination during development or following sciatic nerve crush injury was observed, as determined by quantification of both myelinated and unmyelinated nerve fiber densities, myelinated axonal diameter and myelin thickness. However, the absence of Schwann cell p75NTR reduced motor nerve conduction velocity after crush injury. Our data indicate that the absence of Schwann cell p75NTR expression in vivo is not critical for axonal regrowth or remyelination following sciatic nerve crush injury, but does play a key role in functional recovery. Overall, this represents the first step in redefining the role of p75NTR in the peripheral nervous system, suggesting that the Schwann cell-axon unit functions as a syncytium, with the previous published involvement of p75NTR in remyelination most likely depending on axonal/neuronal p75NTR and/or mutual glial-axonal interactions.
Keywords: Schwann cells; myelination; nerve injury; p75NTR; regeneration.
Figures
Similar articles
-
The neurotrophin receptor p75NTR in Schwann cells is implicated in remyelination and motor recovery after peripheral nerve injury.Glia. 2007 Aug 15;55(11):1199-208. doi: 10.1002/glia.20533. Glia. 2007. PMID: 17600367
-
Schwann cell p75NTR prevents spontaneous sensory reinnervation of the adult spinal cord.Brain. 2010 Feb;133(Pt 2):421-32. doi: 10.1093/brain/awp316. Epub 2010 Jan 3. Brain. 2010. PMID: 20047901
-
Knockout of p75(NTR) impairs re-myelination of injured sciatic nerve in mice.J Neurochem. 2006 Feb;96(3):833-42. doi: 10.1111/j.1471-4159.2005.03564.x. Epub 2005 Nov 29. J Neurochem. 2006. PMID: 16336221
-
p75 Neurotrophin Receptor: A Double-Edged Sword in Pathology and Regeneration of the Central Nervous System.Vet Pathol. 2018 Nov;55(6):786-801. doi: 10.1177/0300985818781930. Epub 2018 Jun 25. Vet Pathol. 2018. PMID: 29940812 Review.
-
[Immortalized adult rodent Schwann cells as useful tools for the study of peripheral nerve regeneration].Rinsho Shinkeigaku. 2013;53(11):1117-9. doi: 10.5692/clinicalneurol.53.1117. Rinsho Shinkeigaku. 2013. PMID: 24291897 Review. Japanese.
Cited by
-
Comparison study of clinicopathological features of cellular schwannoma between retroperitoneum and other sites.Ann Transl Med. 2022 Dec;10(24):1311. doi: 10.21037/atm-22-4979. Ann Transl Med. 2022. PMID: 36660734 Free PMC article.
-
Characterising cellular and molecular features of human peripheral nerve degeneration.Acta Neuropathol Commun. 2020 Apr 17;8(1):51. doi: 10.1186/s40478-020-00921-w. Acta Neuropathol Commun. 2020. PMID: 32303273 Free PMC article.
-
Insights Into the Role and Potential of Schwann Cells for Peripheral Nerve Repair From Studies of Development and Injury.Front Mol Neurosci. 2021 Jan 25;13:608442. doi: 10.3389/fnmol.2020.608442. eCollection 2020. Front Mol Neurosci. 2021. PMID: 33568974 Free PMC article.
-
Schwann cell interactions during the development of the peripheral nervous system.Dev Neurobiol. 2021 Jul;81(5):464-489. doi: 10.1002/dneu.22744. Epub 2020 May 5. Dev Neurobiol. 2021. PMID: 32281247 Free PMC article. Review.
-
A Review on the Role of Endogenous Neurotrophins and Schwann Cells in Axonal Regeneration.J Neuroimmune Pharmacol. 2022 Dec;17(3-4):398-408. doi: 10.1007/s11481-021-10034-3. Epub 2021 Nov 29. J Neuroimmune Pharmacol. 2022. PMID: 34843075 Free PMC article. Review.
References
-
- Bala U., Leong M. P.-Y., Lim C. L., Shahar H. K., Othman F., Lai M.-I., et al. (2018). Defects in nerve conduction velocity and different muscle fibre-type specificity contribute to muscle weakness in Ts1Cje Down syndrome mouse model. PLoS One 13:e0197711. 10.1371/journal.pone.0197711 - DOI - PMC - PubMed
-
- Boyd J. G., Gordon T. (2001). The neurotrophin receptors, trkB and p75, differentially regulate motor axonal regeneration. J. Neurobiol. 49 314–325. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials