

Engineering of L-Plastin Peptide-Loaded Biodegradable Nanoparticles for Sustained Delivery and Suppression of Osteoclast Function In Vitro
- PMID: 31191656
- PMCID: PMC6525930
- DOI: 10.1155/2019/6943986
Engineering of L-Plastin Peptide-Loaded Biodegradable Nanoparticles for Sustained Delivery and Suppression of Osteoclast Function In Vitro
Abstract
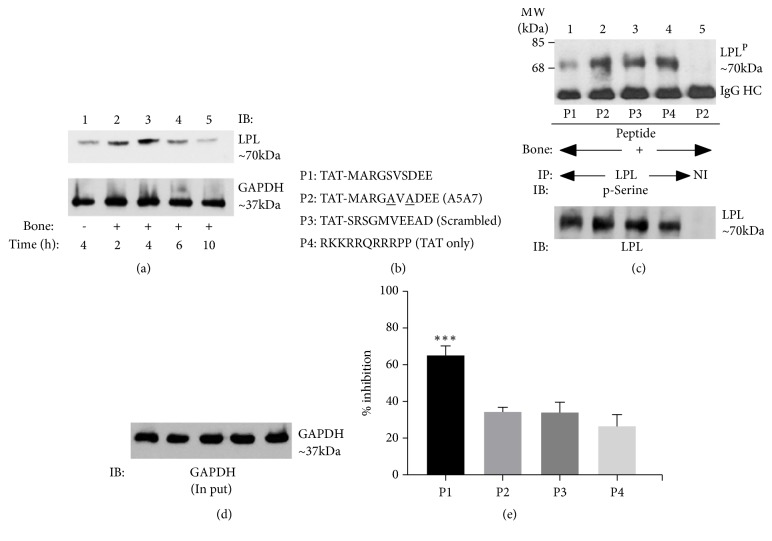
We have recently demonstrated that a small molecular weight amino-terminal peptide of L-plastin (10 amino acids; "MARGSVSDEE") suppressed the phosphorylation of endogenous L-plastin. Therefore, the formation of nascent sealing zones (NSZs) and bone resorption are reduced. The aim of this study was to develop a biodegradable and biocompatible PLGA nanocarrier that could be loaded with the L-plastin peptide of interest and determine the efficacy in vitro in osteoclast cultures. L-plastin MARGSVSDEE (P1) and scrambled control (P3) peptide-loaded PLGA-PEG nanoparticles (NP1 and NP3, respectively) were synthesized by double emulsion technique. The biological effect of nanoparticles on osteoclasts was evaluated by immunoprecipitation, immunoblotting, rhodamine-phalloidin staining of actin filaments, and pit forming assays. Physical characterization of well-dispersed NP1 and NP3 demonstrated ~130-150 nm size, < 0.07 polydispersity index, ~-3 mV ζ-potential, and a sustained release of the peptide for three weeks. Biological characterization in osteoclast cultures demonstrated the following: NP1 significantly reduced (a) endogenous L-plastin phosphorylation; (b) formation of NSZs and sealing rings; (c) resorption. However, the assembly of podosomes which are critical for cell adhesion was not affected. L-plastin peptide-loaded PLGA-PEG nanocarriers have promising potential for the treatment of diseases associated with bone loss. Future studies will use this sustained release of peptide strategy to systematically suppress osteoclast bone resorption activity in vivo in mouse models demonstrating bone loss.
Figures







Similar articles
-
Peptidomimetic inhibitor of L-plastin reduces osteoclastic bone resorption in aging female mice.Bone Res. 2021 Apr 9;9(1):22. doi: 10.1038/s41413-020-00135-9. Bone Res. 2021. PMID: 33837180 Free PMC article.
-
Peptidomimetic inhibitors of L-plastin reduce the resorptive activity of osteoclast but not the bone forming activity of osteoblasts in vitro.PLoS One. 2018 Sep 24;13(9):e0204209. doi: 10.1371/journal.pone.0204209. eCollection 2018. PLoS One. 2018. PMID: 30248139 Free PMC article.
-
L-plastin phosphorylation regulates the early phase of sealing ring formation by actin bundling process in mouse osteoclasts.Exp Cell Res. 2018 Nov 1;372(1):73-82. doi: 10.1016/j.yexcr.2018.09.014. Epub 2018 Sep 21. Exp Cell Res. 2018. PMID: 30244178 Free PMC article.
-
L-Plastin deficiency produces increased trabecular bone due to attenuation of sealing ring formation and osteoclast dysfunction.Bone Res. 2020 Jan 22;8:3. doi: 10.1038/s41413-019-0079-2. eCollection 2020. Bone Res. 2020. PMID: 31993243 Free PMC article.
-
Podosome and sealing zone: specificity of the osteoclast model.Eur J Cell Biol. 2006 Apr;85(3-4):195-202. doi: 10.1016/j.ejcb.2005.09.008. Epub 2005 Oct 24. Eur J Cell Biol. 2006. PMID: 16546562 Review.
Cited by
-
Targeting NF-κB Signaling for Multiple Myeloma.Cancers (Basel). 2020 Aug 6;12(8):2203. doi: 10.3390/cancers12082203. Cancers (Basel). 2020. PMID: 32781681 Free PMC article. Review.
-
C-phycocyanin attenuates RANKL-induced osteoclastogenesis and bone resorption in vitro through inhibiting ROS levels, NFATc1 and NF-κB activation.Sci Rep. 2020 Feb 13;10(1):2513. doi: 10.1038/s41598-020-59363-y. Sci Rep. 2020. PMID: 32054921 Free PMC article.
-
Peptidomimetic inhibitor of L-plastin reduces osteoclastic bone resorption in aging female mice.Bone Res. 2021 Apr 9;9(1):22. doi: 10.1038/s41413-020-00135-9. Bone Res. 2021. PMID: 33837180 Free PMC article.
-
Methylsulfonylmethane increases osteogenesis and regulates the mineralization of the matrix by transglutaminase 2 in SHED cells.PLoS One. 2019 Dec 5;14(12):e0225598. doi: 10.1371/journal.pone.0225598. eCollection 2019. PLoS One. 2019. PMID: 31805069 Free PMC article.
-
The intricate mechanism of PLS3 in bone homeostasis and disease.Front Endocrinol (Lausanne). 2023 Jul 7;14:1168306. doi: 10.3389/fendo.2023.1168306. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37484945 Free PMC article. Review.
References
-
- Teitelbaum S. L., Cao X., Mimura H., Chiba M., Ross F. P. Cellular and molecular mechanisms of bone resorption. Mineral and Electrolyte Metabolism. 1995;21(1-3):193–196. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
