

A new aspect of chronic pain as a lifestyle-related disease
- PMID: 31194049
- PMCID: PMC6550110
- DOI: 10.1016/j.ynpai.2017.04.003
A new aspect of chronic pain as a lifestyle-related disease
Abstract
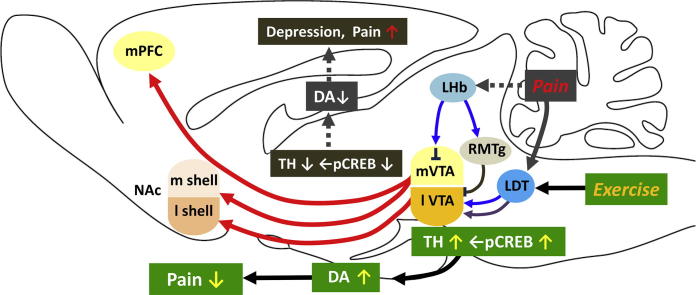
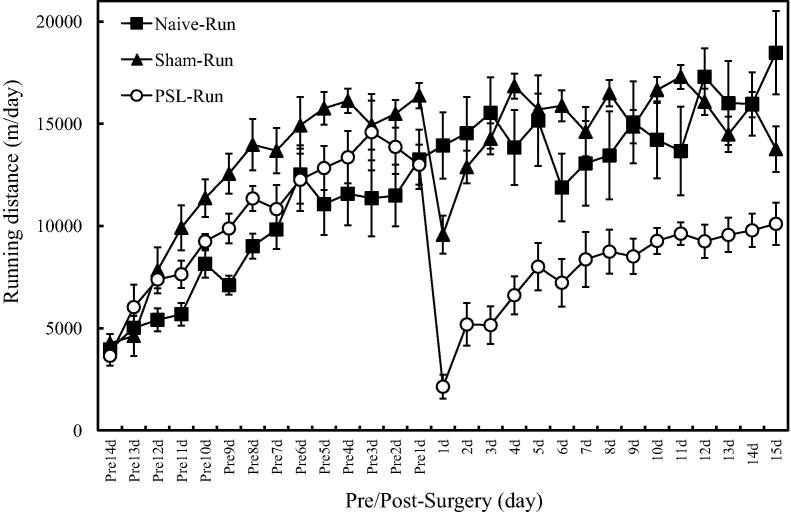
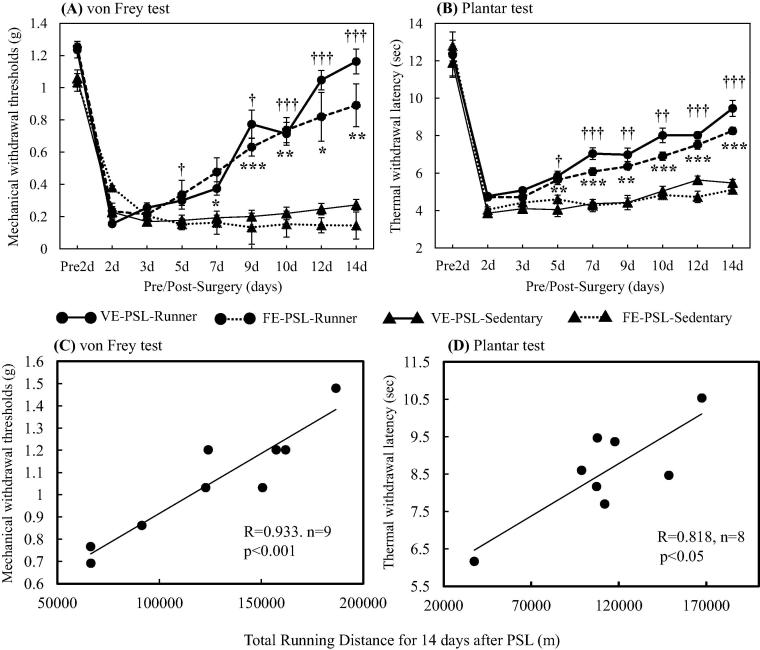
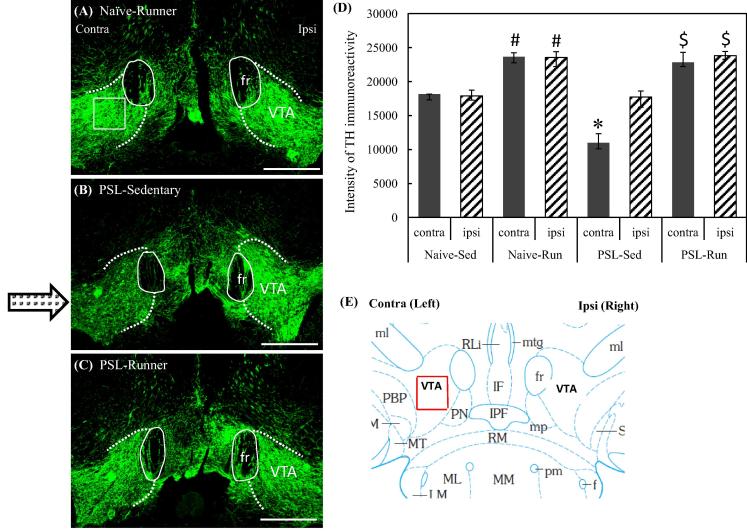
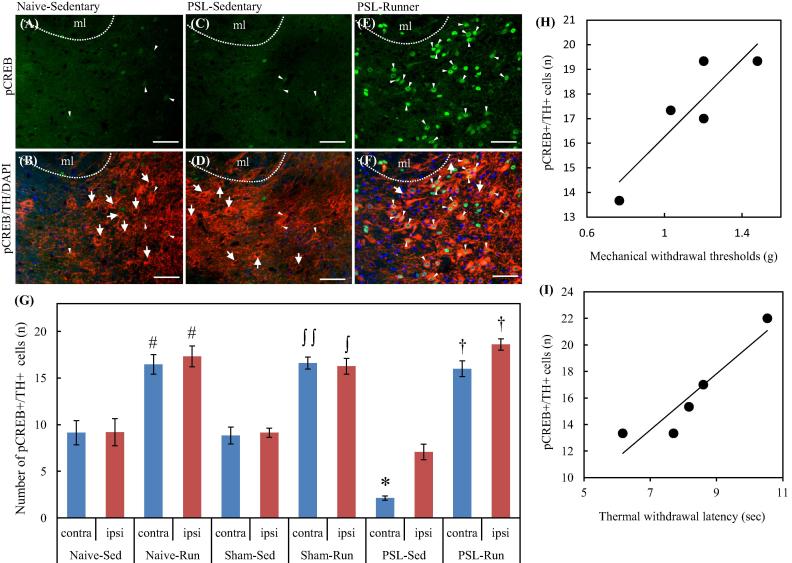
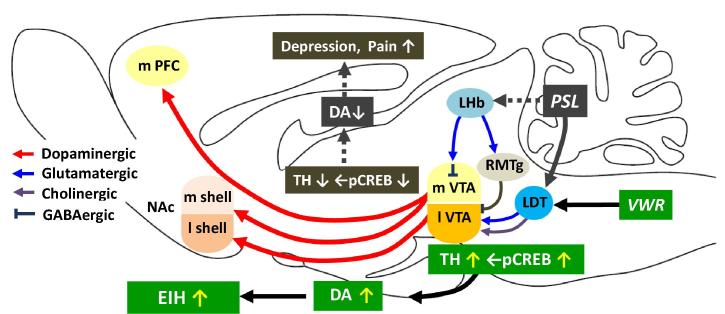
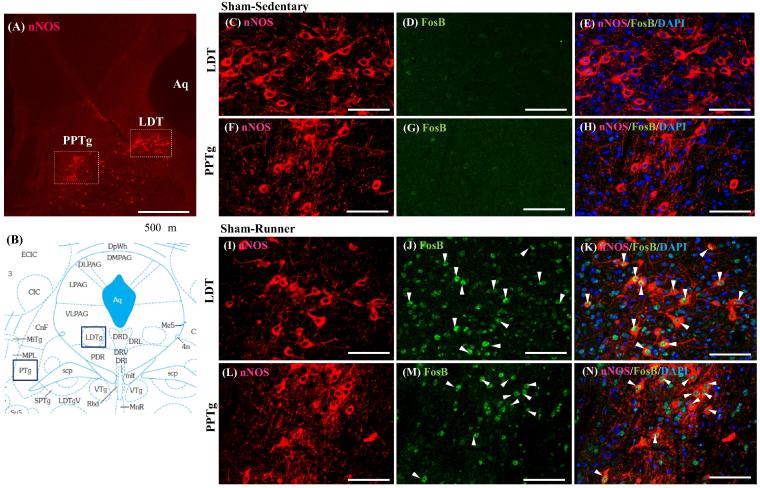
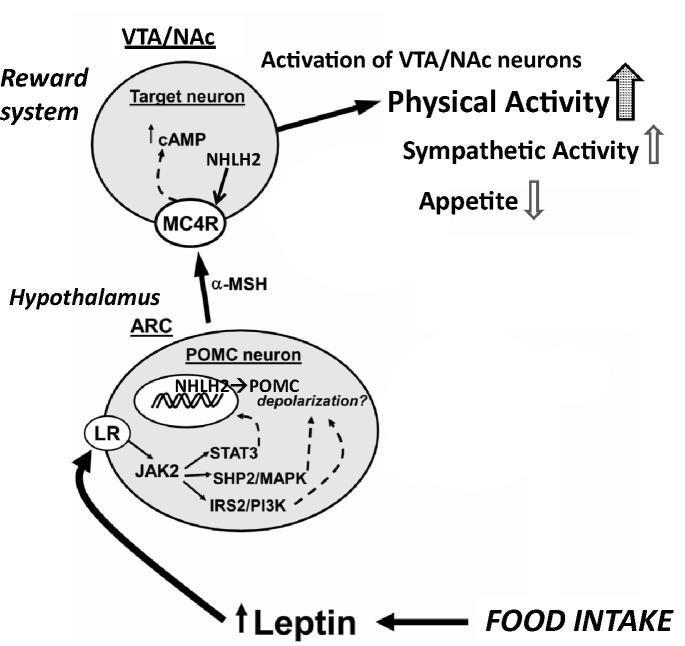
Physical exercise has been established as a low-cost, safe, and effective way to manage chronic intractable pain. We investigated the underlying mechanisms of exercise-induced hypoalgesia (EIH) using a mouse model of neuropathic pain (NPP). Epigenetic changes in activated microglia and maintained GABA synthesis in the spinal dorsal horn may contribute to EIH. Voluntary exercise (VE), a strong reward for animals, also induced EIH, which may be due in part to the activation of dopamine (DA) neurons in the ventral tegmental area (VTA). VE increases the expression of pCREB in dopaminergic neurons in the VTA, which would enhance dopamine production, and thereby contributes to the activation of the mesolimbic reward system in NPP model mice. We demonstrated that neurons in the laterodorsal tegmental and pedunculopontine tegmental nuclei, a major input source of rewarding stimuli to the VTA, were activated by exercise. Chronic pain is at least partly attributed to sedentary and inactive lifestyle as indicated by the Fear-avoidance model. Therefore, chronic pain could be recognized as a lifestyle-related disease. Physical activity/inactivity may be determined by genetic/epigenetic and neural factors encoded in our brain. The hypothalamus and reward system is closely related in the axis of food intake, energy metabolism and physical activity. Understanding the interactions between the mesolimbic DA system and the hypothalamus that sense and regulate energy balance is thus of significant importance. For example, proopiomelanocortin neurons and melanocortin 4 receptors may play a role in connecting these two systems. Therefore, in a certain sense, chronic pain and obesity may share common behavioral and neural pathology, i.e. physical inactivity, as a result of inactivation of the mesolimbic DA system. Exercise and increasing physical activity in daily life may be important in treating and preventing chronic pain, a life-style related disease.
Keywords: CBP, chronic low back pain; Chronic pain; DA, dopamine; Dopamine; Exercise-induced hypoalgesia; FM, fibromyalgia; GABA, gamma-aminobutyric acid; HDAC, histone deacetylase; LDT, laterodorsal tegmental nucleus; LH, lateral hypothalamus; LHb, lateral habenula; Laterodorsal tegmental nucleus; NAc, nucleus accumbens; NPP, neuropathic pain; PPTg, pedunculopontine tegmental nucleus; PSL, partial sciatic nerve ligation; Physical activity/inactivity; RMTg, rostromedial tegmental nucleus; TH, tyrosine hydroxylase; TMD, temporomandibular disorder; VTA, ventral tegmental area; VWR, voluntary wheel running; Ventral tegmental area; delta FosB, delta FBJ murine osteosarcoma viral; mPFC, medial prefrontal cortex; pCREB, phosphorylated cyclic AMP response element-binding protein.
Figures

Similar articles
-
Activation of mesolimbic reward system via laterodorsal tegmental nucleus and hypothalamus in exercise-induced hypoalgesia.Sci Rep. 2018 Aug 1;8(1):11540. doi: 10.1038/s41598-018-29915-4. Sci Rep. 2018. PMID: 30069057 Free PMC article.
-
Disinhibition of Mesolimbic Dopamine Circuit by the Lateral Hypothalamus Regulates Pain Sensation.J Neurosci. 2023 Jun 14;43(24):4525-4540. doi: 10.1523/JNEUROSCI.2298-22.2023. Epub 2023 May 15. J Neurosci. 2023. PMID: 37188517 Free PMC article.
-
Reward and aversion in a heterogeneous midbrain dopamine system.Neuropharmacology. 2014 Jan;76 Pt B(0 0):351-9. doi: 10.1016/j.neuropharm.2013.03.019. Epub 2013 Apr 8. Neuropharmacology. 2014. PMID: 23578393 Free PMC article. Review.
-
Opioid-induced rewards, locomotion, and dopamine activation: A proposed model for control by mesopontine and rostromedial tegmental neurons.Neurosci Biobehav Rev. 2017 Dec;83:72-82. doi: 10.1016/j.neubiorev.2017.09.022. Epub 2017 Sep 23. Neurosci Biobehav Rev. 2017. PMID: 28951251 Free PMC article. Review.
-
Involvement of mesolimbic dopaminergic network in neuropathic pain relief by treadmill exercise: A study for specific neural control with Gi-DREADD in mice.Mol Pain. 2016 Dec 1;12:1744806916681567. doi: 10.1177/1744806916681567. Print 2016. Mol Pain. 2016. PMID: 27909152 Free PMC article.
Cited by
-
Is exercise therapy the first-line treatment for chronic pain?Neurobiol Pain. 2024 Mar 26;15:100154. doi: 10.1016/j.ynpai.2024.100154. eCollection 2024 Jan-Jun. Neurobiol Pain. 2024. PMID: 38881820 Free PMC article. No abstract available.
-
Plastic changes in amygdala subregions by voluntary running contribute to exercise-induced hypoalgesia in neuropathic pain model mice.Mol Pain. 2020 Jan-Dec;16:1744806920971377. doi: 10.1177/1744806920971377. Mol Pain. 2020. PMID: 33297861 Free PMC article.
-
Eating habits and the desire to eat healthier among patients with chronic pain: a registry-based study.Sci Rep. 2024 Feb 27;14(1):4705. doi: 10.1038/s41598-024-55449-z. Sci Rep. 2024. PMID: 38409442 Free PMC article.
-
Tumor suppression and improvement in immune systems by specific activation of dopamine D1-receptor-expressing neurons in the nucleus accumbens.Mol Brain. 2022 Feb 16;15(1):17. doi: 10.1186/s13041-022-00902-1. Mol Brain. 2022. PMID: 35172858 Free PMC article.
-
Serotonergic Neurotransmission System Modulator, Vortioxetine, and Dopaminergic D2/D3 Receptor Agonist, Ropinirole, Attenuate Fibromyalgia-Like Symptoms in Mice.Molecules. 2021 Apr 20;26(8):2398. doi: 10.3390/molecules26082398. Molecules. 2021. PMID: 33924258 Free PMC article.
References
-
- Almeida C., DeMaman A., Kusuda R., Cadetti F., Ravanelli M.I., Queiroz A.L., Sousa T.A., Zanon S., Silveira L.R., Lucas G. Exercise therapy normalizes BDNF upregulation and glial hyperactivity in a mouse model of neuropathic pain. Pain. 2015;156:504–513. - PubMed
-
- Altier N., Stewart J. Dopamine receptor antagonists in the nucleus accumbens attenuate analgesia induced by ventral tegmental area substance P or morphine and by nucleus accumbens amphetamine. J. Pharmacol. Exp. Ther. 1998;285:208–215. - PubMed
-
- Bobinski F., Ferreira T.A., Córdova M.M., Dombrowski P.A., da Cunha C., Santo C.C., Poli A., Pires R.G., Martins-Silva C., Sluka K.A., Santos A.R. Role of brainstem serotonin in analgesia produced by low-intensity exercise on neuropathic pain after sciatic nerve injury in mice. Pain. 2015;156:2595–2606. - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous
