

Two factor authentication: Asf1 mediates crosstalk between H3 K14 and K56 acetylation
- PMID: 31194870
- PMCID: PMC6698667
- DOI: 10.1093/nar/gkz508
Two factor authentication: Asf1 mediates crosstalk between H3 K14 and K56 acetylation
Abstract
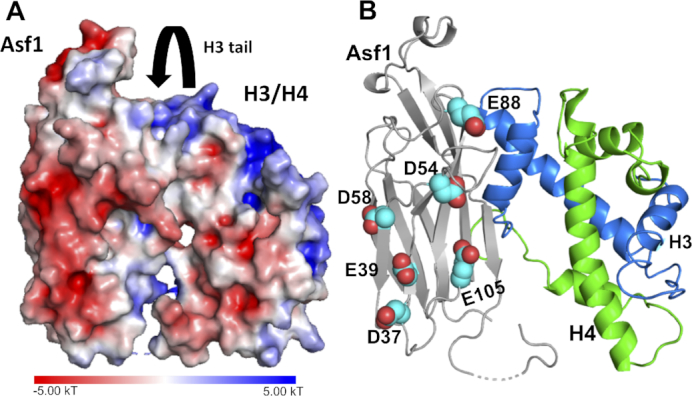
The ability of histone chaperone Anti-silencing factor 1 (Asf1) to direct acetylation of lysine 56 of histone H3 (H3K56ac) represents an important regulatory step in genome replication and DNA repair. In Saccharomyces cerevisiae, Asf1 interacts functionally with a second chaperone, Vps75, and the lysine acetyltransferase (KAT) Rtt109. Both Asf1 and Vps75 can increase the specificity of histone acetylation by Rtt109, but neither alter selectivity. However, changes in acetylation selectivity have been observed in histones extracted from cells, which contain a plethora of post-translational modifications. In the present study, we use a series of singly acetylated histones to test the hypothesis that histone pre-acetylation and histone chaperones function together to drive preferential acetylation of H3K56. We show that pre-acetylated H3K14ac/H4 functions with Asf1 to drive specific acetylation of H3K56 by Rtt109-Vps75. Additionally, we identified an exosite containing an acidic patch in Asf1 and show that mutations to this region alter Asf1-mediated crosstalk that changes Rtt109-Vps75 selectivity. Our proposed mechanism suggests that Gcn5 acetylates H3K14, recruiting remodeler complexes, allowing for the Asf1-H3K14ac/H4 complex to be acetylated at H3K56 by Rtt109-Vps75. This mechanism explains the conflicting biochemical data and the genetic links between Rtt109, Vps75, Gcn5 and Asf1 in the acetylation of H3K56.
Published by Oxford University Press on behalf of Nucleic Acids Research 2019.
Figures







Similar articles
-
Utilizing targeted mass spectrometry to demonstrate Asf1-dependent increases in residue specificity for Rtt109-Vps75 mediated histone acetylation.PLoS One. 2015 Mar 17;10(3):e0118516. doi: 10.1371/journal.pone.0118516. eCollection 2015. PLoS One. 2015. PMID: 25781956 Free PMC article.
-
Chaperone-mediated acetylation of histones by Rtt109 identified by quantitative proteomics.J Proteomics. 2013 Apr 9;81:80-90. doi: 10.1016/j.jprot.2012.09.026. Epub 2012 Oct 2. J Proteomics. 2013. PMID: 23036725 Free PMC article.
-
Structure of the Rtt109-AcCoA/Vps75 complex and implications for chaperone-mediated histone acetylation.Structure. 2011 Feb 9;19(2):221-31. doi: 10.1016/j.str.2010.12.012. Epub 2011 Jan 20. Structure. 2011. PMID: 21256037 Free PMC article.
-
Understanding histone acetyltransferase Rtt109 structure and function: how many chaperones does it take?Curr Opin Struct Biol. 2011 Dec;21(6):728-34. doi: 10.1016/j.sbi.2011.09.005. Epub 2011 Oct 23. Curr Opin Struct Biol. 2011. PMID: 22023828 Free PMC article. Review.
-
Histone-modifying enzymes, histone modifications and histone chaperones in nucleosome assembly: Lessons learned from Rtt109 histone acetyltransferases.Crit Rev Biochem Mol Biol. 2015 Jan-Feb;50(1):31-53. doi: 10.3109/10409238.2014.978975. Epub 2014 Nov 3. Crit Rev Biochem Mol Biol. 2015. PMID: 25365782 Free PMC article. Review.
Cited by
-
Functional interplay of histone lysine 2-hydroxyisobutyrylation and acetylation in Arabidopsis under dark-induced starvation.Nucleic Acids Res. 2021 Jul 21;49(13):7347-7360. doi: 10.1093/nar/gkab536. Nucleic Acids Res. 2021. PMID: 34165567 Free PMC article.
-
Androgens drive SLC1A5-dependent metabolic reprogramming in polycystic ovary syndrome.Nat Commun. 2025 Aug 15;16(1):7611. doi: 10.1038/s41467-025-62951-z. Nat Commun. 2025. PMID: 40817119 Free PMC article.
-
Mapping the residue specificities of epigenome enzymes by yeast surface display.Cell Chem Biol. 2021 Dec 16;28(12):1772-1779.e4. doi: 10.1016/j.chembiol.2021.05.022. Epub 2021 Jun 28. Cell Chem Biol. 2021. PMID: 34186032 Free PMC article.
-
Histone Acetyltransferase Rtt109 Regulates Development, Morphogenesis, and Citrinin Biosynthesis in Monascus purpureus.J Fungi (Basel). 2023 Apr 29;9(5):530. doi: 10.3390/jof9050530. J Fungi (Basel). 2023. PMID: 37233241 Free PMC article.
-
DDK/Hsk1 phosphorylates and targets fission yeast histone deacetylase Hst4 for degradation to stabilize stalled DNA replication forks.Elife. 2021 Oct 5;10:e70787. doi: 10.7554/eLife.70787. Elife. 2021. PMID: 34608864 Free PMC article.
References
-
- Lee D.Y., Hayes J.J., Pruss D., Wolffe A.P.. A positive role for histone acetylation in transcription factor access to nucleosomal DNA. Cell. 1993; 72:73–84. - PubMed
-
- Han J., Zhou H., Horazdovsky B., Zhang K., Xu R.-M., Zhang Z.. Rtt109 acetylates histone H3 lysine 56 and functions in DNA replication. Science. 2007; 315:653–655. - PubMed
-
- Masumoto H., Hawke D., Kobayashi R., Verreault A.. A role for cell-cycle-regulated histone H3 lysine 56 acetylation in the DNA damage response. Nature. 2005; 436:294–298. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
