

Current status and applications of genome-scale metabolic models
- PMID: 31196170
- PMCID: PMC6567666
- DOI: 10.1186/s13059-019-1730-3
Current status and applications of genome-scale metabolic models
Abstract
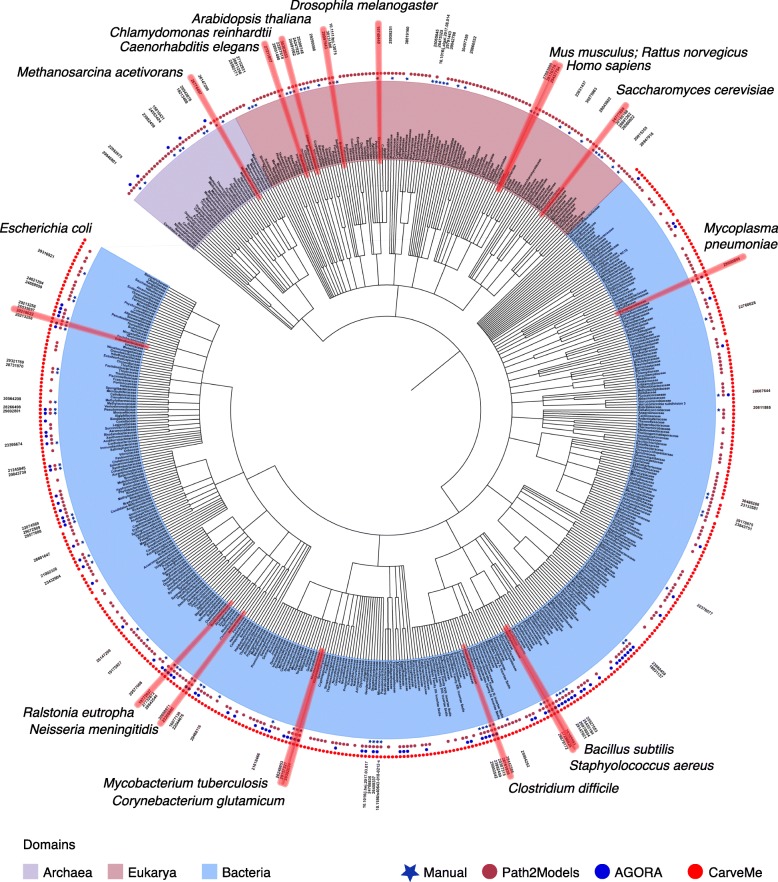
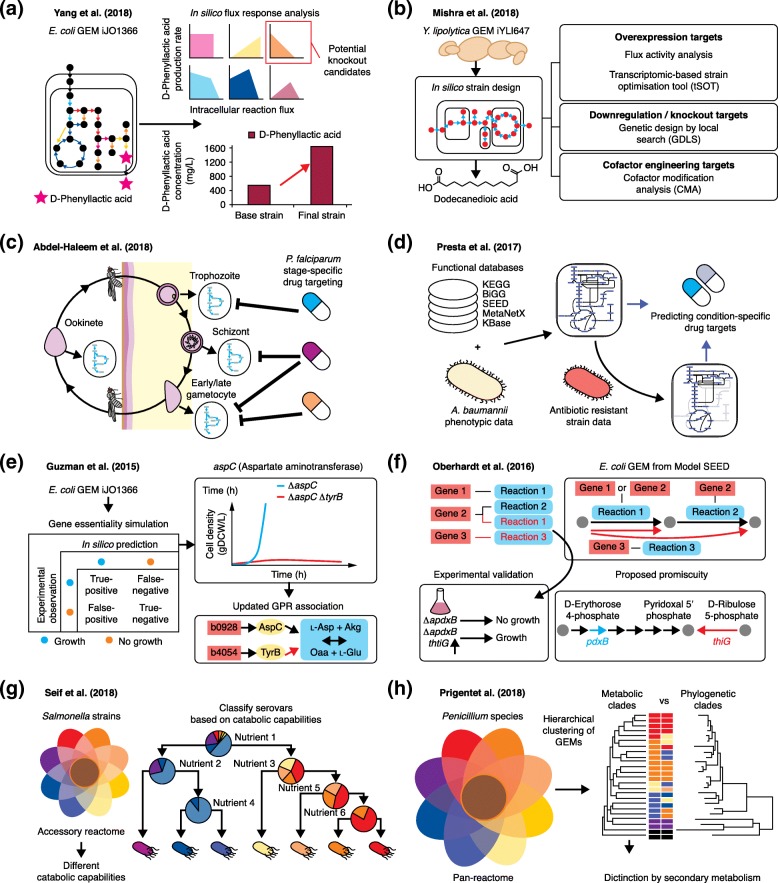
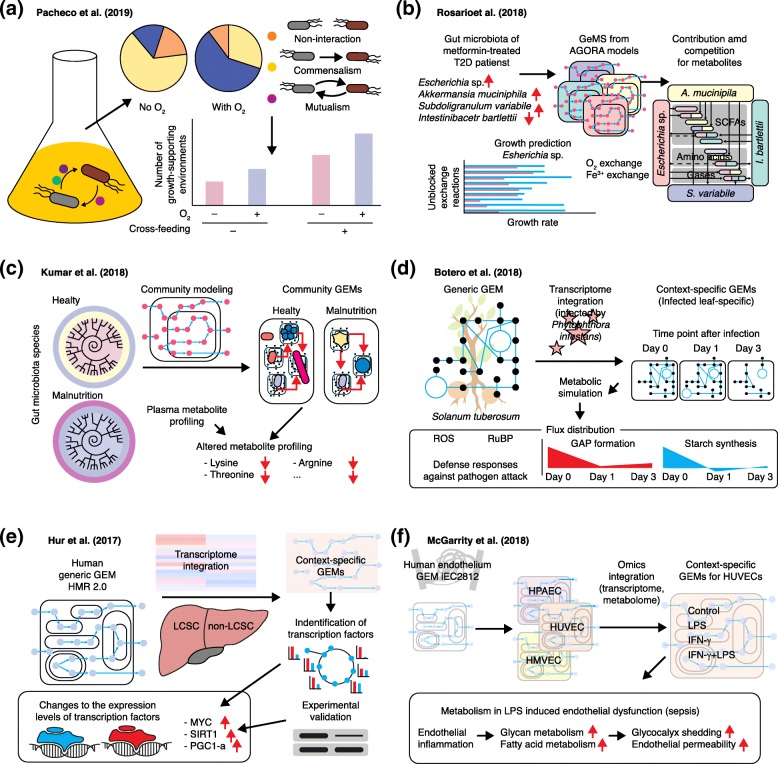
Genome-scale metabolic models (GEMs) computationally describe gene-protein-reaction associations for entire metabolic genes in an organism, and can be simulated to predict metabolic fluxes for various systems-level metabolic studies. Since the first GEM for Haemophilus influenzae was reported in 1999, advances have been made to develop and simulate GEMs for an increasing number of organisms across bacteria, archaea, and eukarya. Here, we review current reconstructed GEMs and discuss their applications, including strain development for chemicals and materials production, drug targeting in pathogens, prediction of enzyme functions, pan-reactome analysis, modeling interactions among multiple cells or organisms, and understanding human diseases.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
