

Low-Frequency Oscillations Code Speech during Verbal Working Memory
- PMID: 31196933
- PMCID: PMC6697399
- DOI: 10.1523/JNEUROSCI.0018-19.2019
Low-Frequency Oscillations Code Speech during Verbal Working Memory
Abstract
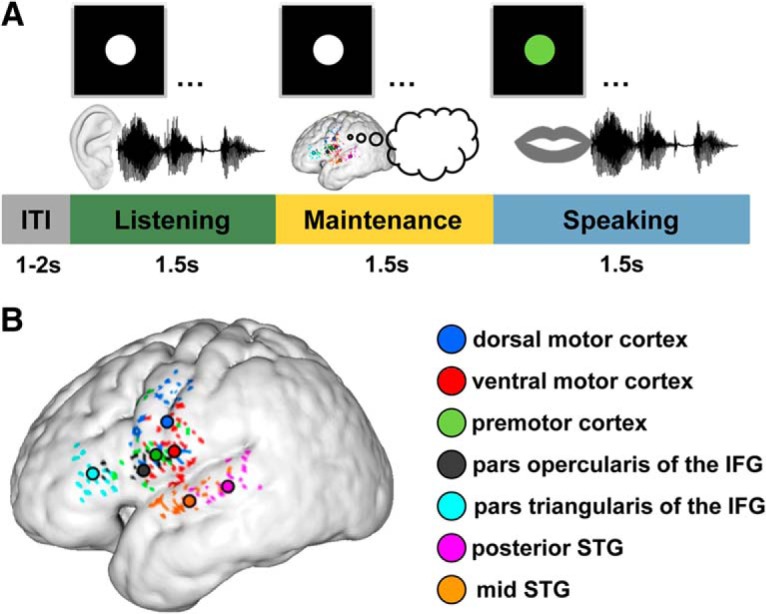
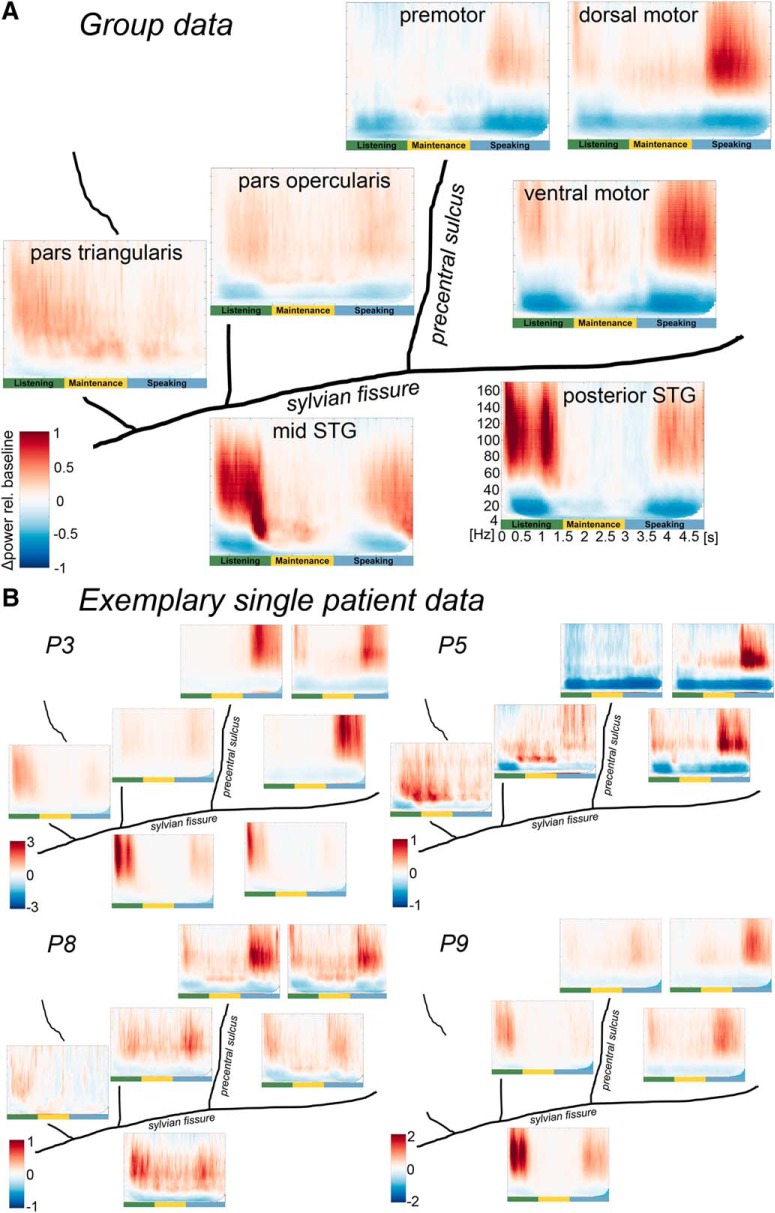
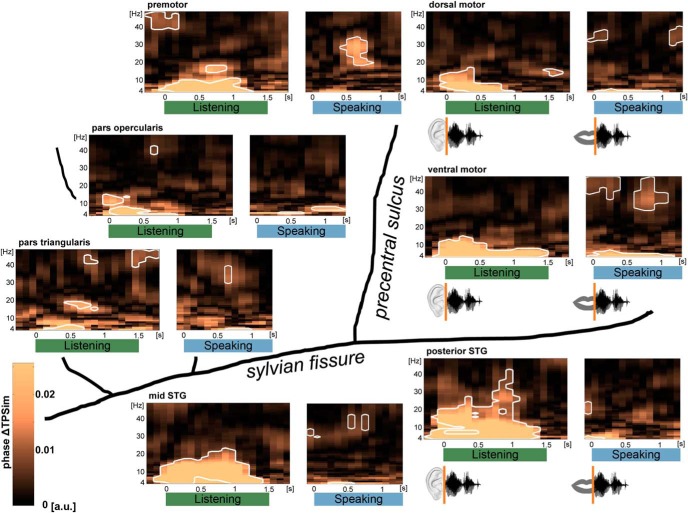
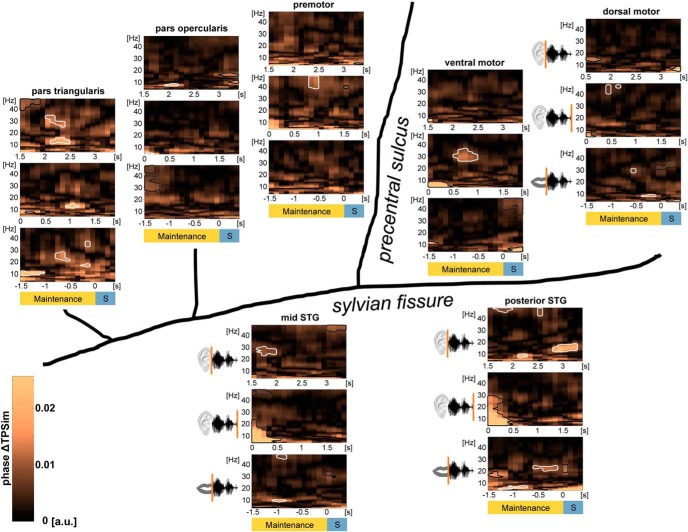
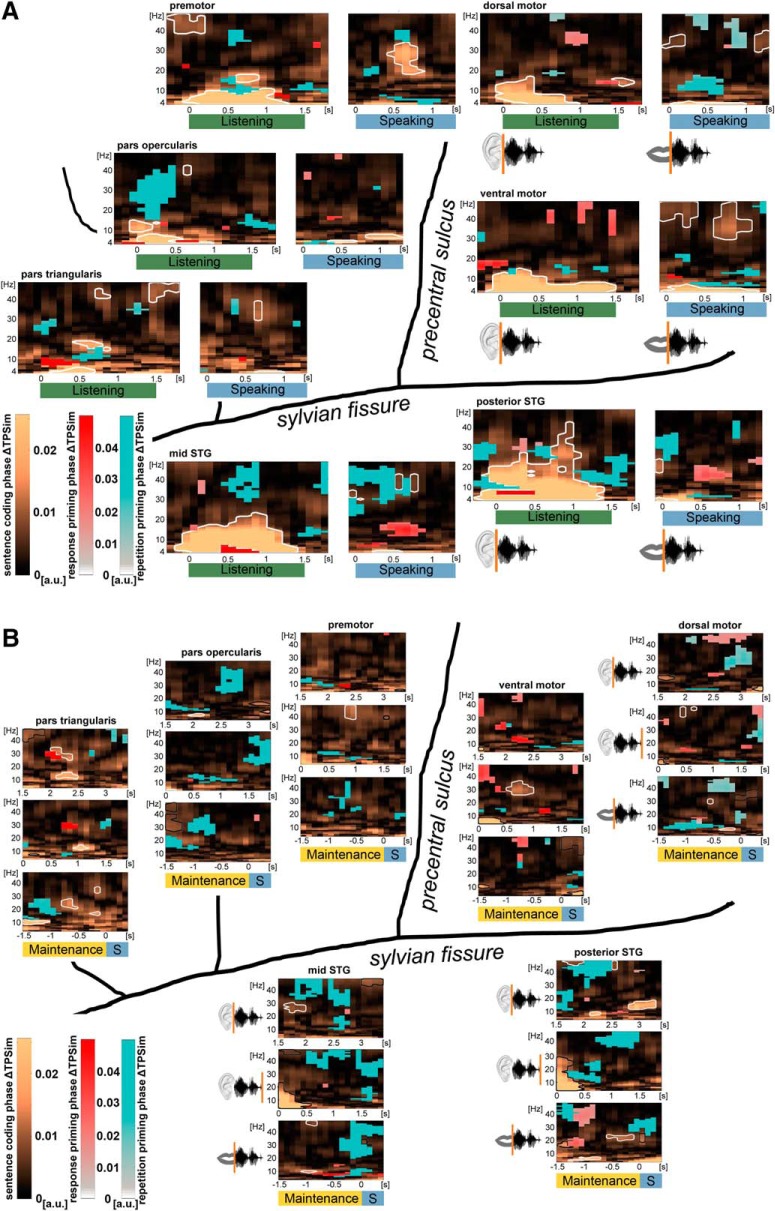
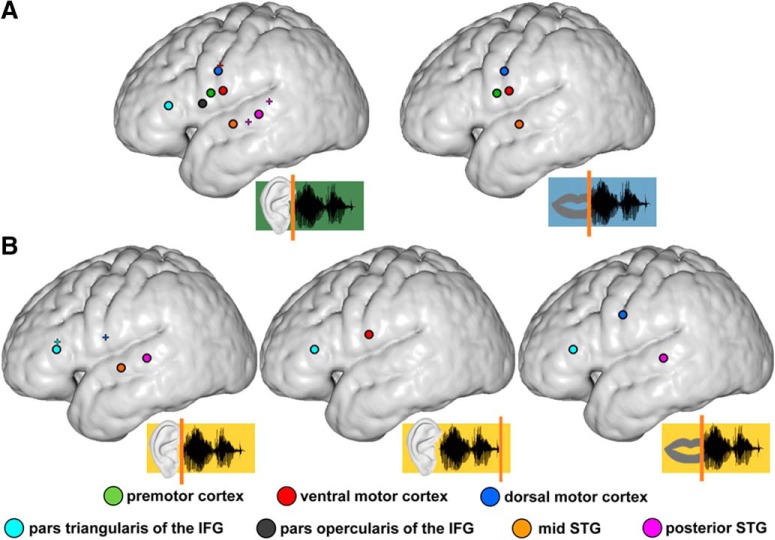
The way the human brain represents speech in memory is still unknown. An obvious characteristic of speech is its evolvement over time. During speech processing, neural oscillations are modulated by the temporal properties of the acoustic speech signal, but also acquired knowledge on the temporal structure of language influences speech perception-related brain activity. This suggests that speech could be represented in the temporal domain, a form of representation that the brain also uses to encode autobiographic memories. Empirical evidence for such a memory code is lacking. We investigated the nature of speech memory representations using direct cortical recordings in the left perisylvian cortex during delayed sentence reproduction in female and male patients undergoing awake tumor surgery. Our results reveal that the brain endogenously represents speech in the temporal domain. Temporal pattern similarity analyses revealed that the phase of frontotemporal low-frequency oscillations, primarily in the beta range, represents sentence identity in working memory. The positive relationship between beta power during working memory and task performance suggests that working memory representations benefit from increased phase separation.SIGNIFICANCE STATEMENT Memory is an endogenous source of information based on experience. While neural oscillations encode autobiographic memories in the temporal domain, little is known on their contribution to memory representations of human speech. Our electrocortical recordings in participants who maintain sentences in memory identify the phase of left frontotemporal beta oscillations as the most prominent information carrier of sentence identity. These observations provide evidence for a theoretical model on speech memory representations and explain why interfering with beta oscillations in the left inferior frontal cortex diminishes verbal working memory capacity. The lack of sentence identity coding at the syllabic rate suggests that sentences are represented in memory in a more abstract form compared with speech coding during speech perception and production.
Keywords: electrocorticography; memory representations; sentence repetition; speech perception; speech production; temporal pattern similarity.
Copyright © 2019 the authors.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources