

SFI1 promotes centriole duplication by recruiting USP9X to stabilize the microcephaly protein STIL
- PMID: 31197030
- PMCID: PMC6605807
- DOI: 10.1083/jcb.201803041
SFI1 promotes centriole duplication by recruiting USP9X to stabilize the microcephaly protein STIL
Abstract
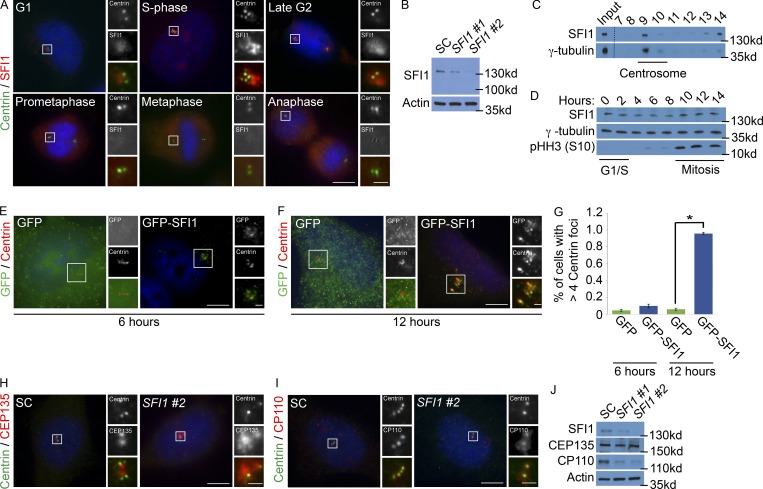
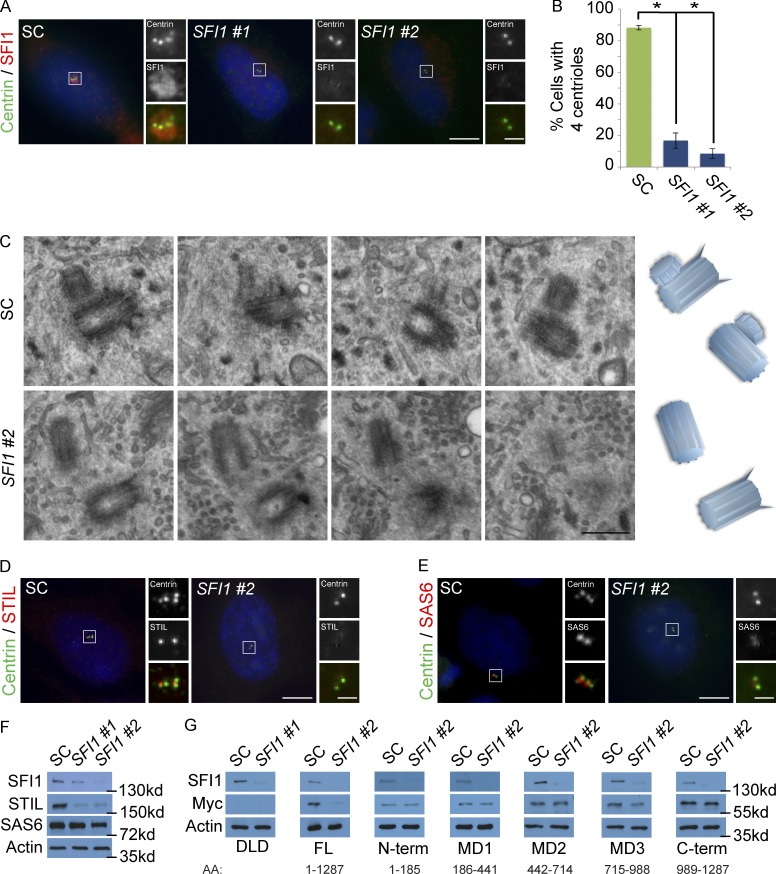
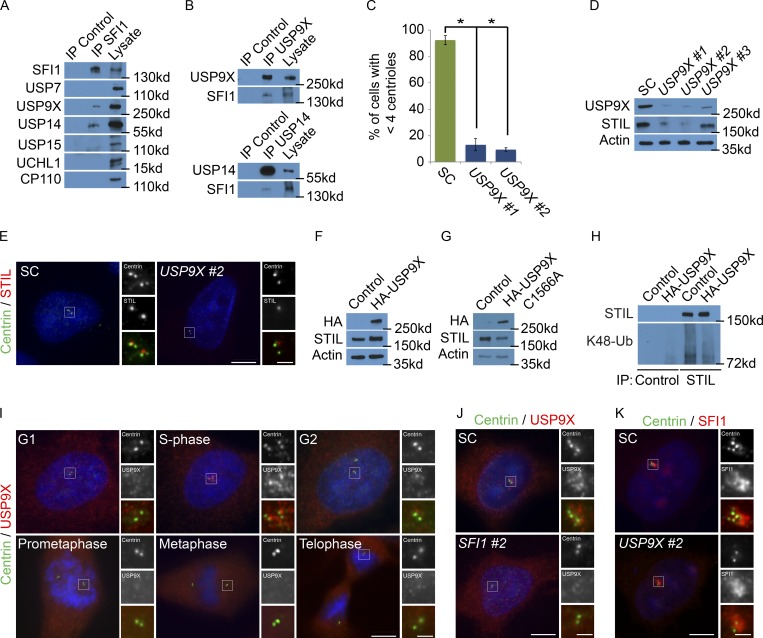
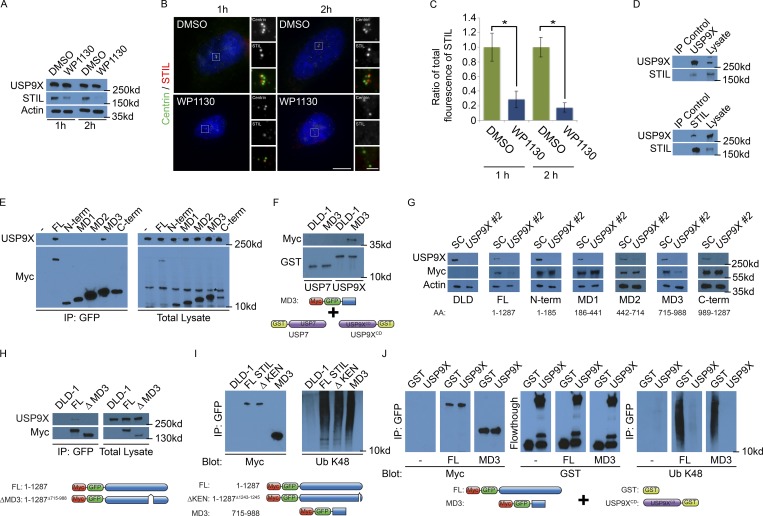
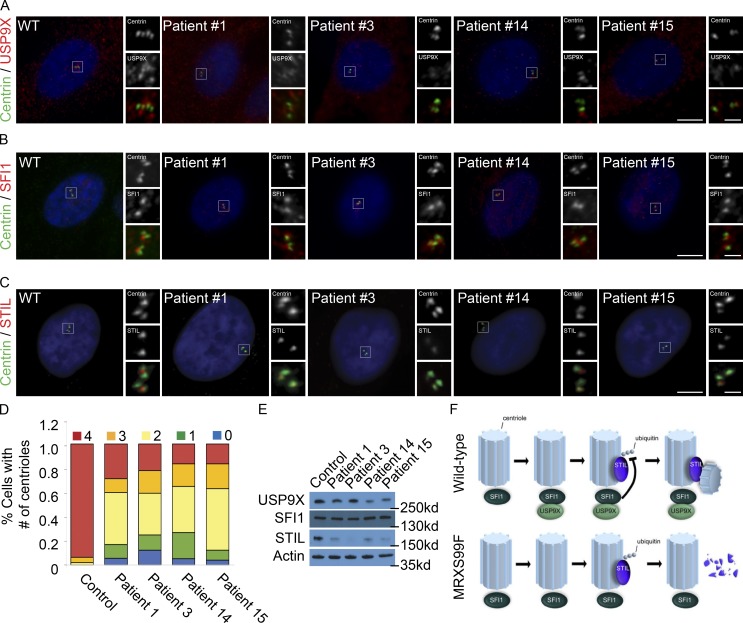
In mammals, centrioles participate in brain development, and human mutations affecting centriole duplication cause microcephaly. Here, we identify a role for the mammalian homologue of yeast SFI1, involved in the duplication of the yeast spindle pole body, as a critical regulator of centriole duplication in mammalian cells. Mammalian SFI1 interacts with USP9X, a deubiquitylase associated with human syndromic mental retardation. SFI1 localizes USP9X to the centrosome during S phase to deubiquitylate STIL, a critical regulator of centriole duplication. USP9X-mediated deubiquitylation protects STIL from degradation. Consistent with a role for USP9X in stabilizing STIL, cells from patients with USP9X loss-of-function mutations have reduced STIL levels. Together, these results demonstrate that SFI1 is a centrosomal protein that localizes USP9X to the centrosome to stabilize STIL and promote centriole duplication. We propose that the USP9X protection of STIL to facilitate centriole duplication underlies roles of both proteins in human neurodevelopment.
© 2019 Kodani et al.
Figures
Similar articles
-
STIL is required for centriole duplication in human cells.J Cell Sci. 2012 Mar 1;125(Pt 5):1353-62. doi: 10.1242/jcs.104109. Epub 2012 Feb 20. J Cell Sci. 2012. PMID: 22349705
-
The human microcephaly protein STIL interacts with CPAP and is required for procentriole formation.EMBO J. 2011 Oct 21;30(23):4790-804. doi: 10.1038/emboj.2011.378. EMBO J. 2011. PMID: 22020124 Free PMC article.
-
STIL microcephaly mutations interfere with APC/C-mediated degradation and cause centriole amplification.Curr Biol. 2014 Feb 17;24(4):351-60. doi: 10.1016/j.cub.2013.12.016. Epub 2014 Jan 30. Curr Biol. 2014. PMID: 24485834
-
The centrosome duplication cycle in health and disease.FEBS Lett. 2014 Aug 1;588(15):2366-72. doi: 10.1016/j.febslet.2014.06.030. Epub 2014 Jun 18. FEBS Lett. 2014. PMID: 24951839 Review.
-
Ubiquitin signaling in the control of centriole duplication.FEBS J. 2022 Aug;289(16):4830-4849. doi: 10.1111/febs.16069. Epub 2021 Jun 29. FEBS J. 2022. PMID: 34115927 Review.
Cited by
-
USP9X Is Required to Maintain Cell Survival in Response to High-LET Radiation.Front Oncol. 2021 Jul 1;11:671431. doi: 10.3389/fonc.2021.671431. eCollection 2021. Front Oncol. 2021. PMID: 34277417 Free PMC article.
-
Ubiquitin system mutations in neurological diseases.Trends Biochem Sci. 2024 Oct;49(10):875-887. doi: 10.1016/j.tibs.2024.06.011. Epub 2024 Jul 6. Trends Biochem Sci. 2024. PMID: 38972780 Review.
-
Deubiquitylases in developmental ubiquitin signaling and congenital diseases.Cell Death Differ. 2021 Feb;28(2):538-556. doi: 10.1038/s41418-020-00697-5. Epub 2020 Dec 17. Cell Death Differ. 2021. PMID: 33335288 Free PMC article. Review.
-
Case report: NUT carcinoma with MXI1::NUTM1 fusion characterized by abdominopelvic lesions and ovarian masses in a middle-aged female.Front Oncol. 2023 Jan 11;12:1091877. doi: 10.3389/fonc.2022.1091877. eCollection 2022. Front Oncol. 2023. PMID: 36741693 Free PMC article.
-
Tetrahymena Poc5 is a transient basal body component that is important for basal body maturation.J Cell Sci. 2020 Jun 4;133(11):jcs240838. doi: 10.1242/jcs.240838. J Cell Sci. 2020. PMID: 32350068 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
