

Emergence of plasmid stability under non-selective conditions maintains antibiotic resistance
- PMID: 31197163
- PMCID: PMC6565834
- DOI: 10.1038/s41467-019-10600-7
Emergence of plasmid stability under non-selective conditions maintains antibiotic resistance
Abstract
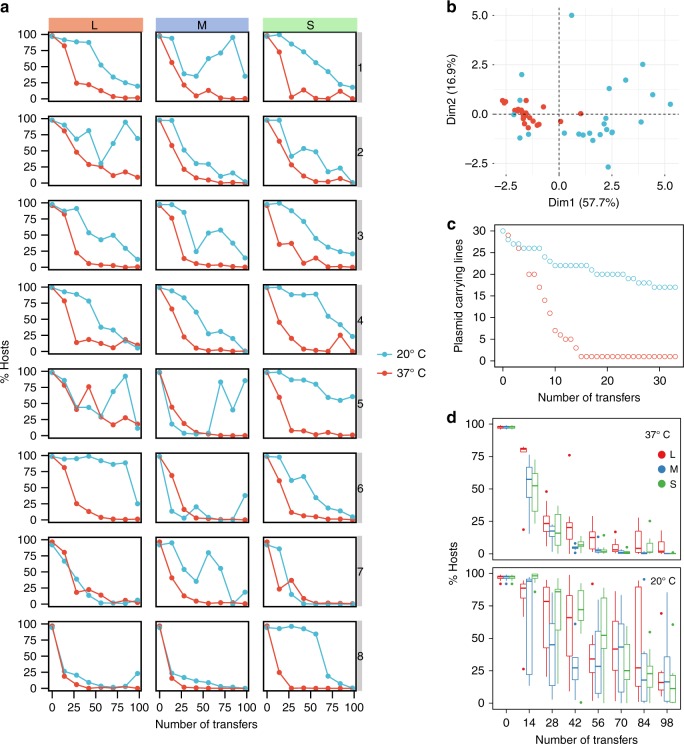
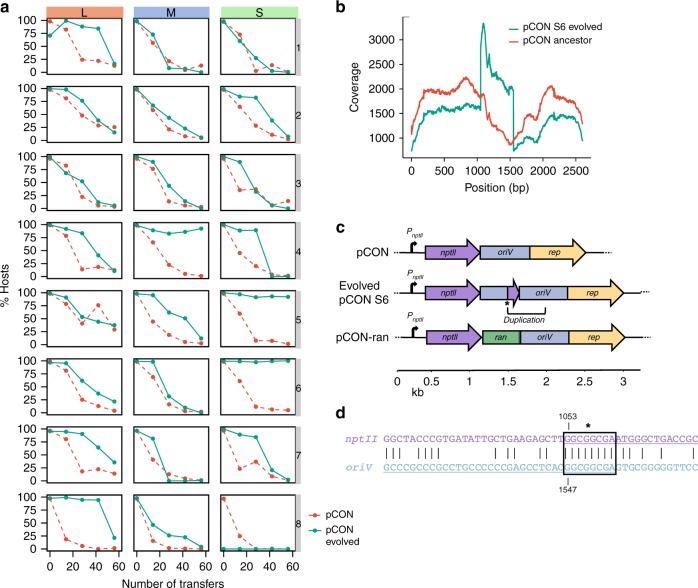
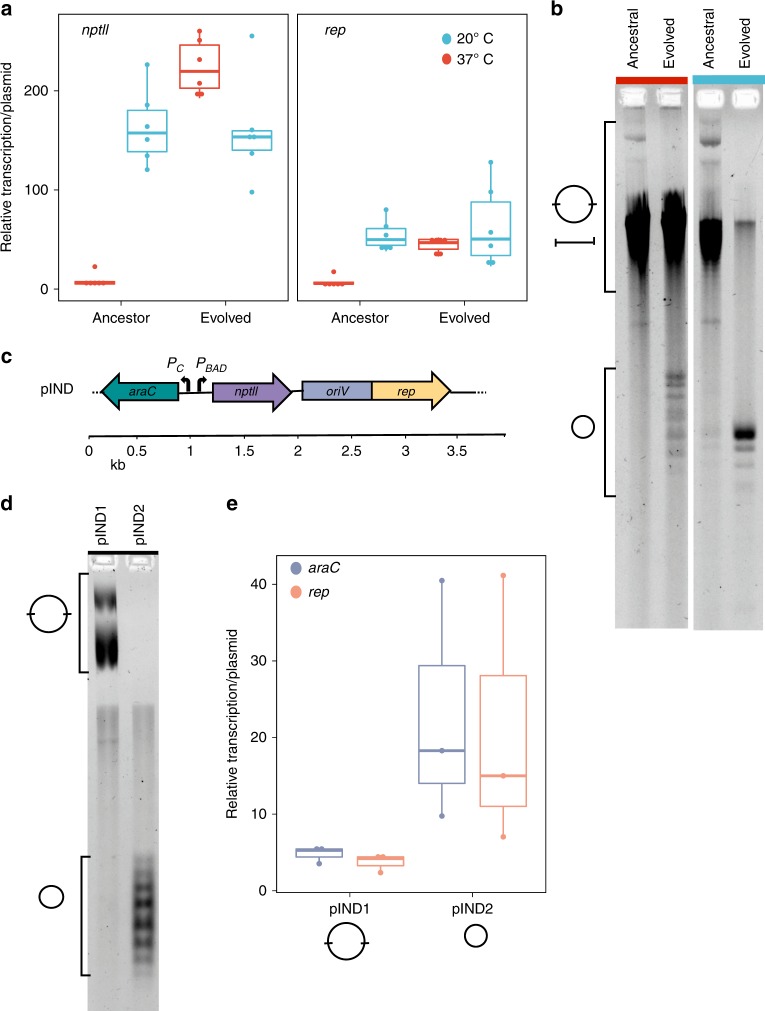
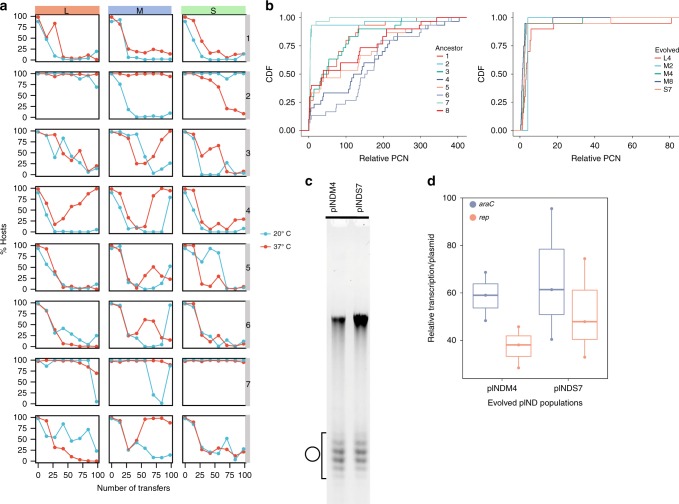
Plasmid acquisition is an important mechanism of rapid adaptation and niche expansion in prokaryotes. Positive selection for plasmid-coded functions is a major driver of plasmid evolution, while plasmids that do not confer a selective advantage are considered costly and expected to go extinct. Yet, plasmids are ubiquitous in nature, and their persistence remains an evolutionary paradox. Here, we demonstrate that non-mobile plasmids persist over evolutionary timescales without selection for the plasmid function. Evolving a minimal plasmid encoding for antibiotics resistance in Escherichia coli, we discover that plasmid stability emerges in the absence of antibiotics and that plasmid loss is determined by transcription-replication conflicts. We further find that environmental conditions modulate these conflicts and plasmid persistence. Silencing the transcription of the resistance gene results in stable plasmids that become fixed in the population. Evolution of plasmid stability under non-selective conditions provides an evolutionary explanation for the ubiquity of plasmids in nature.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Antibiotics Interfere with the Evolution of Plasmid Stability.Curr Biol. 2020 Oct 5;30(19):3841-3847.e4. doi: 10.1016/j.cub.2020.07.019. Epub 2020 Aug 13. Curr Biol. 2020. PMID: 32795438
-
Survival and Evolution of a Large Multidrug Resistance Plasmid in New Clinical Bacterial Hosts.Mol Biol Evol. 2016 Nov;33(11):2860-2873. doi: 10.1093/molbev/msw163. Epub 2016 Aug 8. Mol Biol Evol. 2016. PMID: 27501945 Free PMC article.
-
Increased copy number couples the evolution of plasmid horizontal transmission and plasmid-encoded antibiotic resistance.Proc Natl Acad Sci U S A. 2021 Aug 3;118(31):e2107818118. doi: 10.1073/pnas.2107818118. Proc Natl Acad Sci U S A. 2021. PMID: 34326267 Free PMC article.
-
Plasmid persistence: costs, benefits, and the plasmid paradox.Can J Microbiol. 2018 May;64(5):293-304. doi: 10.1139/cjm-2017-0609. Epub 2018 Mar 21. Can J Microbiol. 2018. PMID: 29562144 Review.
-
Evolution of Plasmid-Mediated Antibiotic Resistance in the Clinical Context.Trends Microbiol. 2018 Dec;26(12):978-985. doi: 10.1016/j.tim.2018.06.007. Epub 2018 Jul 23. Trends Microbiol. 2018. PMID: 30049587 Review.
Cited by
-
Piggybacking on Niche Adaptation Improves the Maintenance of Multidrug-Resistance Plasmids.Mol Biol Evol. 2021 Jul 29;38(8):3188-3201. doi: 10.1093/molbev/msab091. Mol Biol Evol. 2021. PMID: 33760032 Free PMC article.
-
Genomic epidemiology of emerging ESBL-producing Salmonella Kentucky bla CTX-M-14b in Europe.Emerg Microbes Infect. 2020 Dec;9(1):2124-2135. doi: 10.1080/22221751.2020.1821582. Emerg Microbes Infect. 2020. PMID: 32896234 Free PMC article.
-
Adaptive evolution of plasmid and chromosome contributes to the fitness of a blaNDM-bearing cointegrate plasmid in Escherichia coli.ISME J. 2024 Jan 8;18(1):wrae037. doi: 10.1093/ismejo/wrae037. ISME J. 2024. PMID: 38438143 Free PMC article.
-
Application of antimicrobial, potential hazard and mitigation plans.Environ Res. 2022 Dec;215(Pt 1):114218. doi: 10.1016/j.envres.2022.114218. Epub 2022 Aug 29. Environ Res. 2022. PMID: 36049514 Free PMC article. Review.
-
mMGE: a database for human metagenomic extrachromosomal mobile genetic elements.Nucleic Acids Res. 2021 Jan 8;49(D1):D783-D791. doi: 10.1093/nar/gkaa869. Nucleic Acids Res. 2021. PMID: 33074335 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
