

Timely expression and activation of YAP1 in granulosa cells is essential for ovarian follicle development
- PMID: 31199671
- PMCID: PMC6704445
- DOI: 10.1096/fj.201900179RR
Timely expression and activation of YAP1 in granulosa cells is essential for ovarian follicle development
Abstract
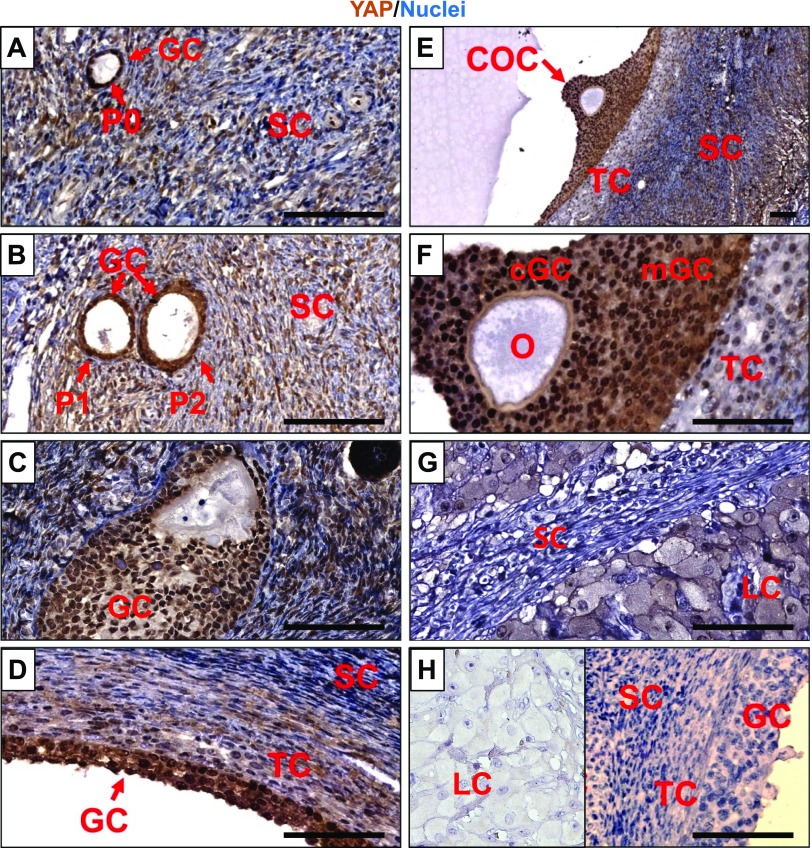
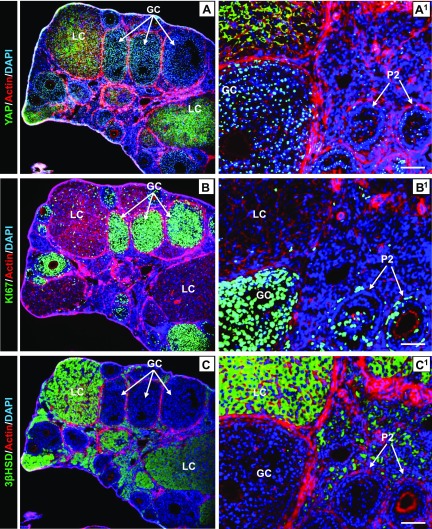
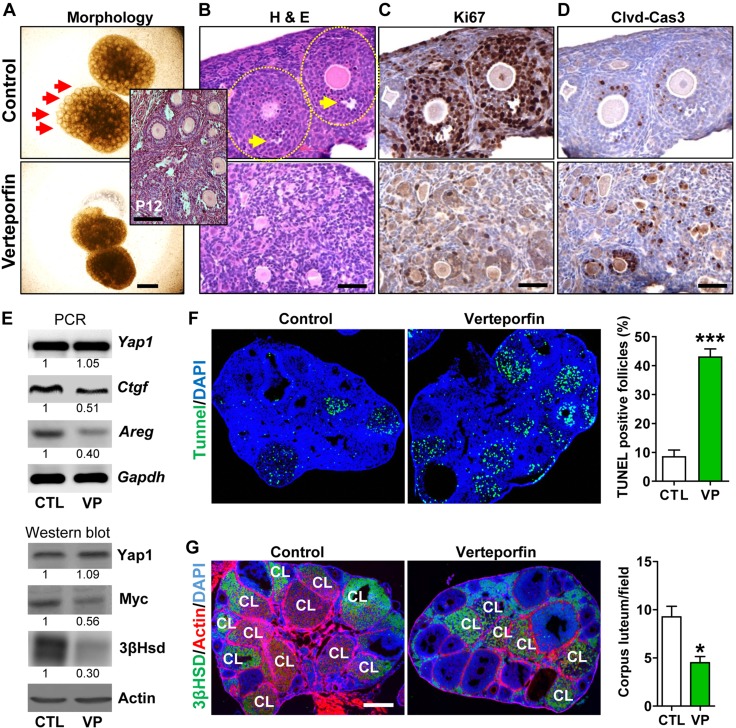
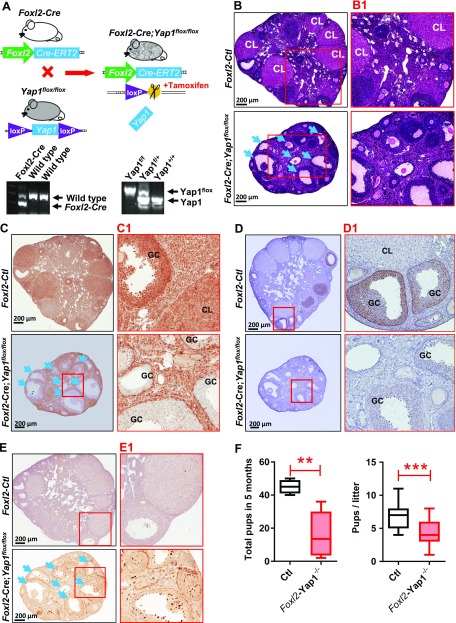
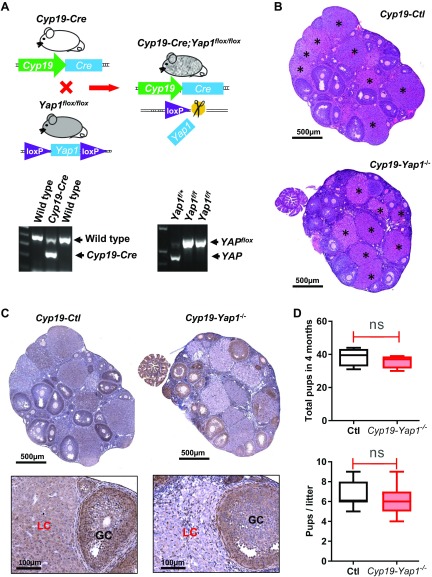
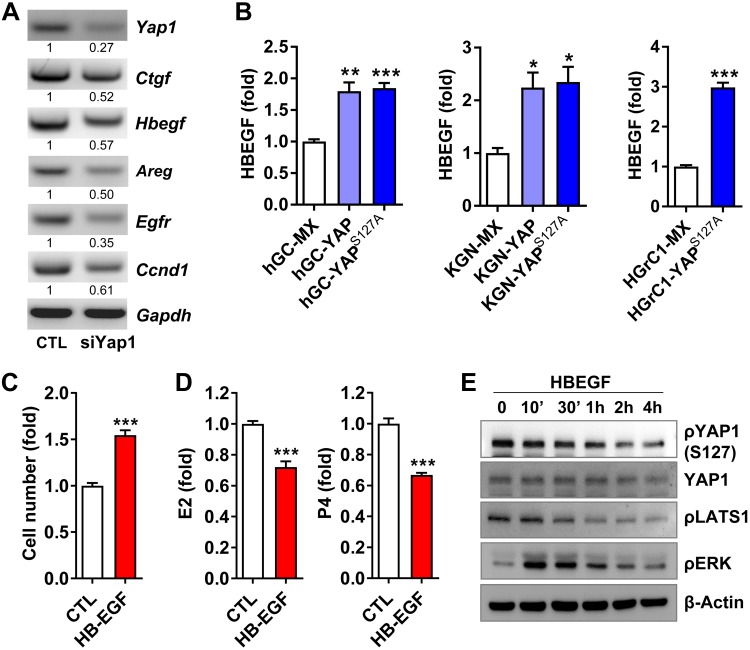
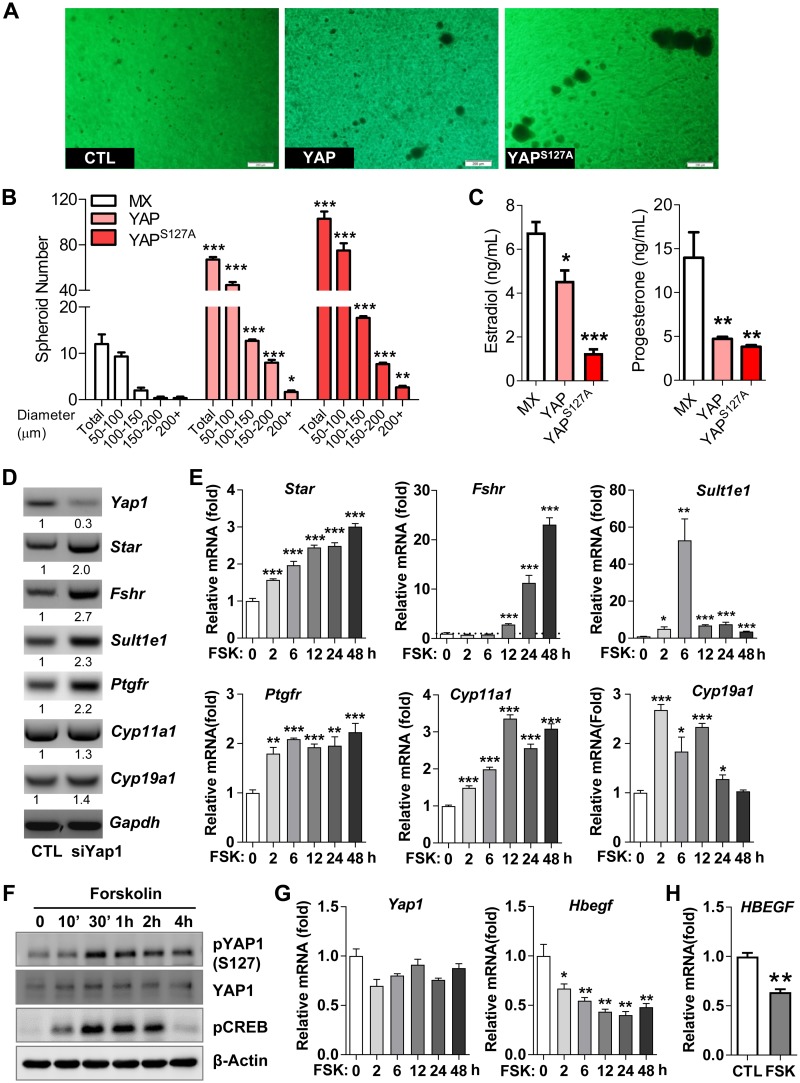
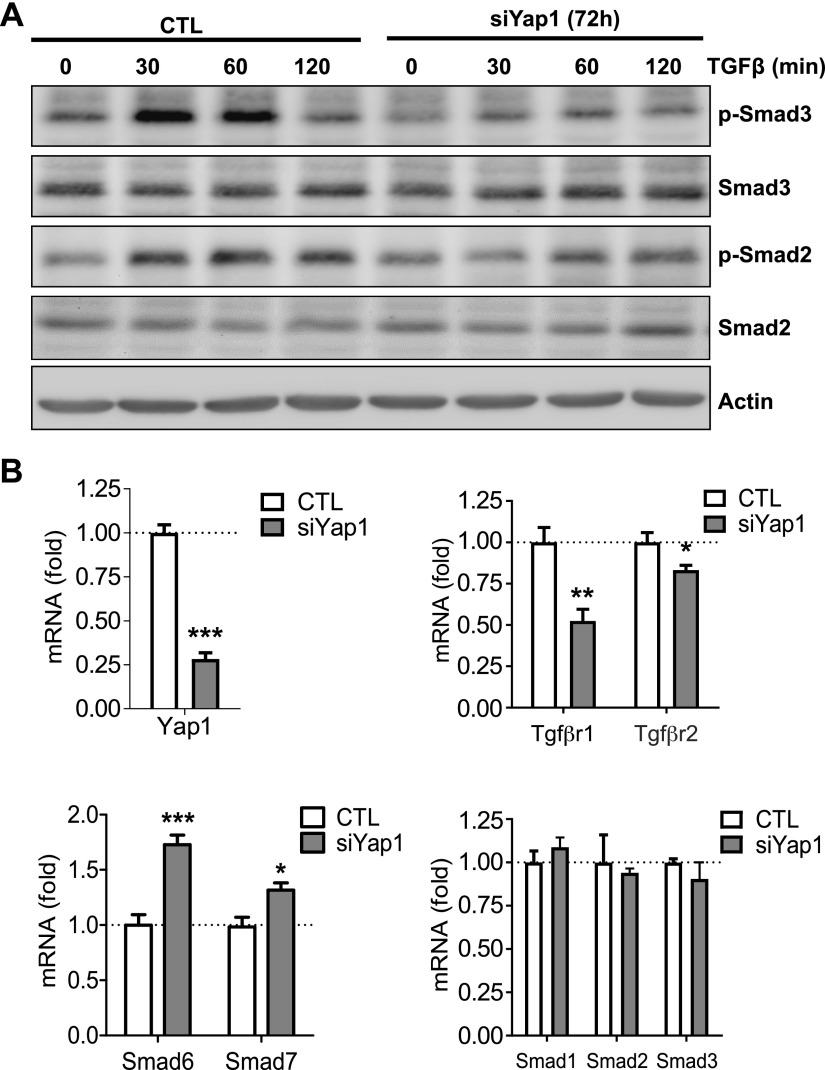
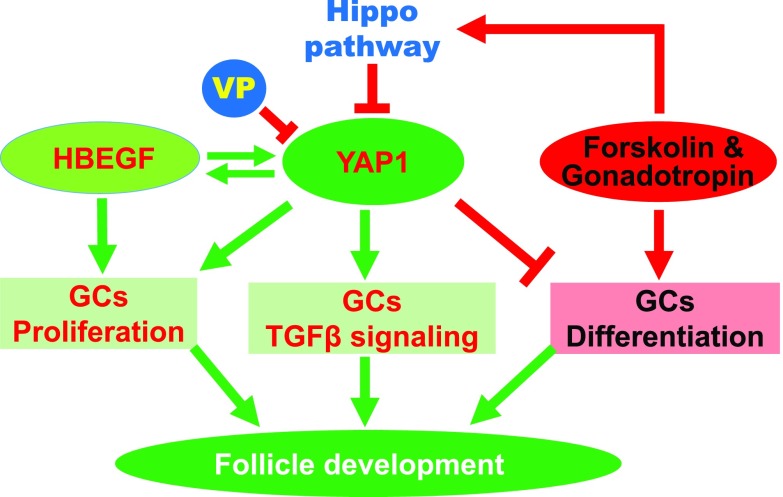
Although the role of the Hippo signaling pathway in development and tumorigenesis has been extensively studied in multiple organs, its role in ovarian follicle development remains largely unknown. Here, we report that Yes-Associated Protein 1 (YAP1), the major effector of Hippo signaling, is spatiotemporally expressed in ovarian granulosa cells and plays a critical role in the regulation of follicle development. We found that the active form of YAP1 (nuclear YAP1) was predominantly expressed in proliferative granulosa cells, whereas the inactive form of YAP1 (cytoplasmic YAP1) was mainly detected in luteal cells (terminally differentiated granulosa cells). Pharmacological inhibition of YAP1 activity disrupted mouse ovarian follicle development in vitro and in vivo. Foxl2 promoter-driven knockout of Yap1 in ovarian granulosa cells resulted in increased apoptosis of granulosa cells, decreased number of corpora lutea, reduced ovarian size, and subfertility in transgenic mice. However, Cyp19a1 promoter-driven knockout of Yap1 in differentiated granulosa cells of preovulatory follicles and luteal cells of corpora lutea had no effect on ovarian morphology and fertility. Mechanistic studies demonstrated that YAP1 interacted with epidermal growth factor receptor and TGF-β signaling pathways to regulate granulosa cell proliferation, differentiation, and survival. Results from this study identify YAP1 as a critical regulator of granulosa cell proliferation and differentiation. Balanced expression and activation of YAP1 is essential for follicle development and successful reproduction. YAP1 is a promising target for treatment of subfertility associated with abnormal granulosa cell function.-Lv, X., He, C., Huang, C., Wang, H., Hua, G., Wang, Z., Zhou, J., Chen, X., Ma, B., Timm, B. K., Maclin, V., Dong, J., Rueda, B. R., Davis, J. S., Wang, C. Timely expression and activation of YAP1 in granulosa cells is essential for ovarian follicle development.
Keywords: Hippo pathway; Steroidogenesis; Yes-Associated Protein 1; fertility.
Conflict of interest statement
The authors thank Eric N. Olson (University of Texas Southwestern Medical Center, Dallas, TX) for providing the Yap1-floxed mouse model. The authors also thank Janice A. Taylor and James R. Talaska of the Advanced Microscopy Core Facility at the University of Nebraska Medical Center for providing assistance with confocal microscopy. This work was supported by a University of Nebraska Medical Center Fellowship; the Eunice Kennedy Shriver National Institute of Child Health and Human Development (5R00HD059985); The National Cancer Institute and U.S. National Institutes of Health (NIH) (1R01CA197976, 1R01CA201500), the Vincent Center for Reproductive Biology, Department of Obstetrics and Gynecology, Massachusetts General Hospital–Harvard Medical School; the Olson Center for Women’s Health at the University of Nebraska Medical Center; The Department of Veteran’s Affairs, and the Colleen’s Dream Foundation. X.L., C. He, and C. Huang are co-first authors. The authors declare no conflicts of interest.
Figures
References
-
- Stocco C., Telleria C., Gibori G. (2007) The molecular control of corpus luteum formation, function, and regression. Endocr. Rev. 28, 117–149 - PubMed
-
- Polesello C., Tapon N. (2007) Salvador-warts-hippo signaling promotes Drosophila posterior follicle cell maturation downstream of notch. Curr. Biol. 17, 1864–1870 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
