

Protein interactions and consensus clustering analysis uncover insights into herpesvirus virion structure and function relationships
- PMID: 31199794
- PMCID: PMC6594648
- DOI: 10.1371/journal.pbio.3000316
Protein interactions and consensus clustering analysis uncover insights into herpesvirus virion structure and function relationships
Abstract
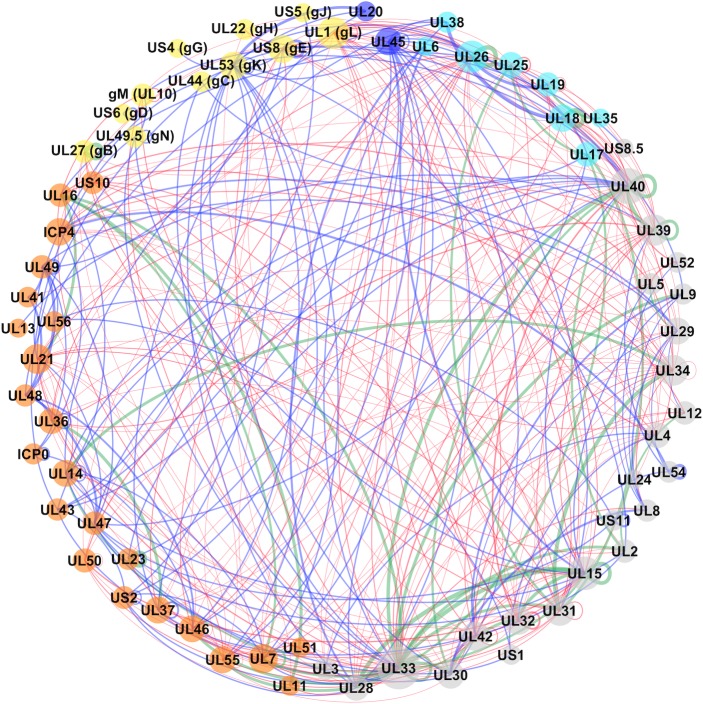
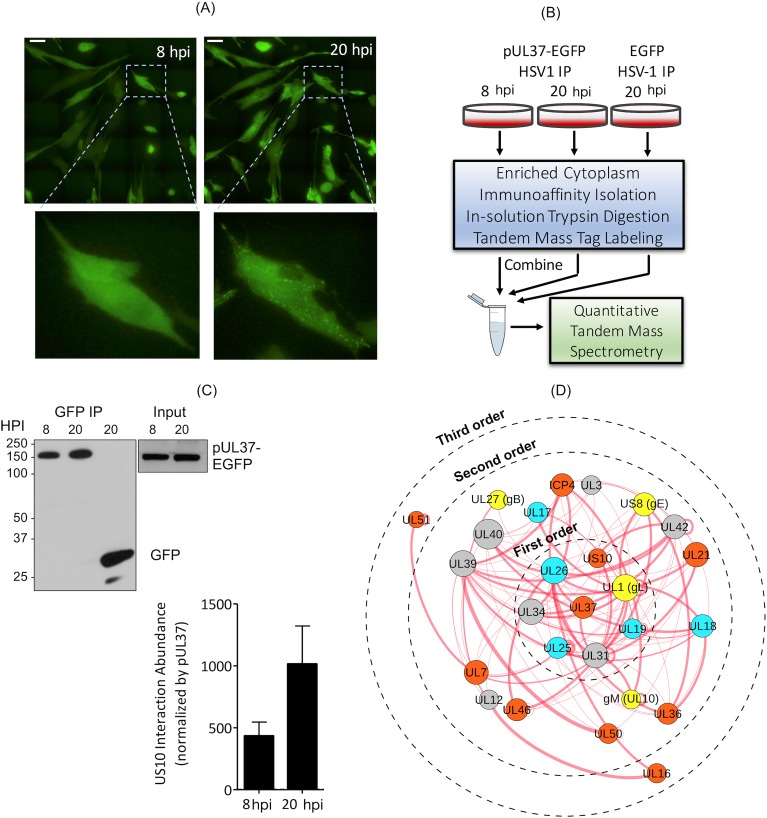
Infections with human herpesviruses are ubiquitous and a public health concern worldwide. Current treatments reduce the severity of some symptoms associated to herpetic infections but neither remove the viral reservoir from the infected host nor protect from the recurrent symptom outbreaks that characterise herpetic infections. The difficulty in therapeutically tackling these viral systems stems in part from their remarkably large proteomes and the complex networks of physical and functional associations that they tailor. This study presents our efforts to unravel the complexity of the interactome of herpes simplex virus type 1 (HSV1), the prototypical herpesvirus species. Inspired by our previous work, we present an improved and more integrative computational pipeline for the protein-protein interaction (PPI) network reconstruction in HSV1, together with a newly developed consensus clustering framework, which allowed us to extend the analysis beyond binary physical interactions and revealed a system-level layout of higher-order functional associations in the virion proteome. Additionally, the analysis provided new functional annotation for the currently undercharacterised protein pUS10. In-depth bioinformatics sequence analysis unravelled structural features in pUS10 reminiscent of those observed in some capsid-associated proteins in tailed bacteriophages, with which herpesviruses are believed to share a common ancestry. Using immunoaffinity purification (IP)-mass spectrometry (MS), we obtained additional support for our bioinformatically predicted interaction between pUS10 and the inner tegument protein pUL37, which binds cytosolic capsids, contributing to initial tegumentation and eventually virion maturation. In summary, this study unveils new, to our knowledge, insights at both the system and molecular levels that can help us better understand the complexity behind herpesvirus infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Cytosolic herpes simplex virus capsids not only require binding inner tegument protein pUL36 but also pUL37 for active transport prior to secondary envelopment.Cell Microbiol. 2013 Feb;15(2):248-69. doi: 10.1111/cmi.12075. Epub 2012 Dec 20. Cell Microbiol. 2013. PMID: 23186167
-
Conserved Tryptophan Motifs in the Large Tegument Protein pUL36 Are Required for Efficient Secondary Envelopment of Herpes Simplex Virus Capsids.J Virol. 2016 May 12;90(11):5368-5383. doi: 10.1128/JVI.03167-15. Print 2016 Jun 1. J Virol. 2016. PMID: 27009950 Free PMC article.
-
Comprehensive Analysis of the Tegument Proteins Involved in Capsid Transport and Virion Morphogenesis of Alpha, Beta and Gamma Herpesviruses.Viruses. 2023 Oct 6;15(10):2058. doi: 10.3390/v15102058. Viruses. 2023. PMID: 37896835 Free PMC article. Review.
-
The C terminus of the large tegument protein pUL36 contains multiple capsid binding sites that function differently during assembly and cell entry of herpes simplex virus.J Virol. 2012 Apr;86(7):3682-700. doi: 10.1128/JVI.06432-11. Epub 2012 Jan 18. J Virol. 2012. PMID: 22258258 Free PMC article.
-
Herpesvirus capsid assembly: insights from structural analysis.Curr Opin Virol. 2011 Aug;1(2):142-9. doi: 10.1016/j.coviro.2011.06.003. Curr Opin Virol. 2011. PMID: 21927635 Free PMC article. Review.
Cited by
-
Conserved Central Intraviral Protein Interactome of the Herpesviridae Family.mSystems. 2019 Oct 1;4(5):e00295-19. doi: 10.1128/mSystems.00295-19. mSystems. 2019. PMID: 31575665 Free PMC article.
-
Functional Domains of the Herpes Simplex Virus Type 1 Tegument Protein pUL37: The Amino Terminus is Dispensable for Virus Replication in Tissue Culture.Viruses. 2019 Sep 14;11(9):853. doi: 10.3390/v11090853. Viruses. 2019. PMID: 31540043 Free PMC article.
-
Alphaherpesvirus glycoprotein E: A review of its interactions with other proteins of the virus and its application in vaccinology.Front Microbiol. 2022 Aug 4;13:970545. doi: 10.3389/fmicb.2022.970545. eCollection 2022. Front Microbiol. 2022. PMID: 35992696 Free PMC article. Review.
-
Temporal dynamics of protein complex formation and dissociation during human cytomegalovirus infection.Nat Commun. 2020 Feb 10;11(1):806. doi: 10.1038/s41467-020-14586-5. Nat Commun. 2020. PMID: 32041945 Free PMC article.
-
A network view of human immune system and virus-human interaction.Front Immunol. 2022 Oct 26;13:997851. doi: 10.3389/fimmu.2022.997851. eCollection 2022. Front Immunol. 2022. PMID: 36389817 Free PMC article.
References
-
- Arvin A, Campadelli-Fiume G, Mocarski E, Moore PS, Roizman B, Whitley R, et al. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis. Cambridge: Cambridge University Press; 2007. - PubMed
-
- Readhead B, Haure-Mirande J-V, Funk CC, Richards MA, Shannon P, Haroutunian V, et al. Multiscale Analysis of Independent Alzheimer’s Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks by Human Herpesvirus. Neuron. Cell Press; 2018;99: 64–82.e7. 10.1016/j.neuron.2018.05.023 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
