

Mitosis-related phosphorylation of the eukaryotic translation suppressor 4E-BP1 and its interaction with eukaryotic translation initiation factor 4E (eIF4E)
- PMID: 31201269
- PMCID: PMC6682726
- DOI: 10.1074/jbc.RA119.008512
Mitosis-related phosphorylation of the eukaryotic translation suppressor 4E-BP1 and its interaction with eukaryotic translation initiation factor 4E (eIF4E)
Erratum in
-
Correction: Mitosis-related phosphorylation of the eukaryotic translation suppressor 4E-BP1 and its interaction with eukaryotic translation initiation factor 4E (eIF4E).J Biol Chem. 2020 Apr 3;295(14):4760. doi: 10.1074/jbc.AAC120.013398. J Biol Chem. 2020. PMID: 32245903 Free PMC article. No abstract available.
Abstract
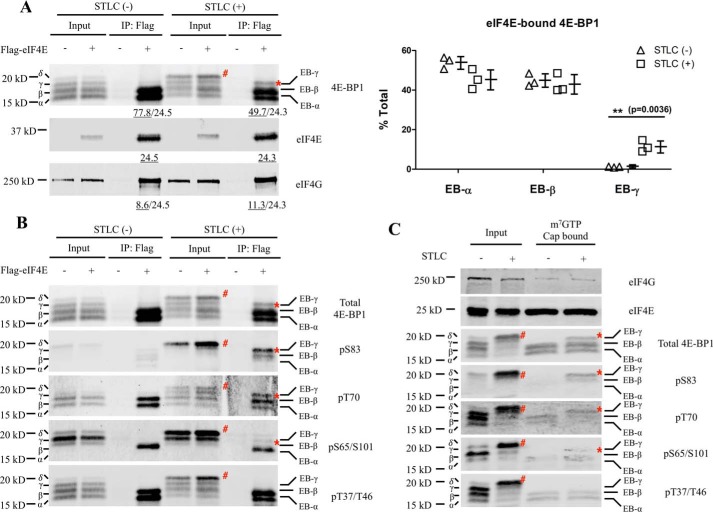
Eukaryotic translation initiation factor 4E (eIF4E)-binding protein 1 (4E-BP1) inhibits cap-dependent translation in eukaryotes by competing with eIF4G for an interaction with eIF4E. Phosphorylation at Ser-83 of 4E-BP1 occurs during mitosis through the activity of cyclin-dependent kinase 1 (CDK1)/cyclin B rather than through canonical mTOR kinase activity. Here, we investigated the interaction of eIF4E with 4E-BP1 or eIF4G during interphase and mitosis. We observed that 4E-BP1 and eIF4G bind eIF4E at similar levels during interphase and mitosis. The most highly phosphorylated mitotic 4E-BP1 isoform (δ) did not interact with eIF4E, whereas a distinct 4E-BP1 phospho-isoform, EB-γ, phosphorylated at Thr-70, Ser-83, and Ser-101, bound to eIF4E during mitosis. Two-dimensional gel electrophoretic analysis corroborated the identity of the phosphorylation marks on the eIF4E-bound 4E-BP1 isoforms and uncovered a population of phosphorylated 4E-BP1 molecules lacking Thr-37/Thr-46-priming phosphorylation. Moreover, proximity ligation assays for phospho-4E-BP1 and eIF4E revealed different in situ interactions during interphase and mitosis. The eIF4E:eIF4G interaction was not inhibited but rather increased in mitotic cells, consistent with active translation initiation during mitosis. Phosphodefective substitution of 4E-BP1 at Ser-83 did not change global translation or individual mRNA translation profiles as measured by single-cell nascent protein synthesis and eIF4G RNA immunoprecipitation sequencing. Mitotic 5'-terminal oligopyrimidine RNA translation was active and, unlike interphase translation, resistant to mTOR inhibition. Our findings reveal the phosphorylation profiles of 4E-BP1 isoforms and their interactions with eIF4E throughout the cell cycle and indicate that 4E-BP1 does not specifically inhibit translation initiation during mitosis.
Keywords: PHAS-I; cell cycle; cyclin-dependent kinase 1 (CDK1); eukaryotic translation initiation; eukaryotic translation initiation factor 4E (eIF4E); eukaryotic translation initiation factor 4E-binding protein 1 (EIF4EBP1); eukaryotic translation initiation factor 4G (eIF4G); mammalian target of rapamycin (mTOR); mitosis; protein phosphorylation.
© 2019 Sun et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures






Similar articles
-
CDK1 substitutes for mTOR kinase to activate mitotic cap-dependent protein translation.Proc Natl Acad Sci U S A. 2015 May 12;112(19):5875-82. doi: 10.1073/pnas.1505787112. Epub 2015 Apr 16. Proc Natl Acad Sci U S A. 2015. PMID: 25883264 Free PMC article.
-
Acute oral leucine administration stimulates protein synthesis during chronic sepsis through enhanced association of eukaryotic initiation factor 4G with eukaryotic initiation factor 4E in rats.J Nutr. 2007 Sep;137(9):2074-9. doi: 10.1093/jn/137.9.2074. J Nutr. 2007. PMID: 17709445
-
New hierarchical phosphorylation pathway of the translational repressor eIF4E-binding protein 1 (4E-BP1) in ischemia-reperfusion stress.J Biol Chem. 2010 Nov 5;285(45):34355-63. doi: 10.1074/jbc.M110.135103. Epub 2010 Aug 24. J Biol Chem. 2010. PMID: 20736160 Free PMC article.
-
4E-BP1, a multifactor regulated multifunctional protein.Cell Cycle. 2016;15(6):781-6. doi: 10.1080/15384101.2016.1151581. Cell Cycle. 2016. PMID: 26901143 Free PMC article. Review.
-
The mTOR/4E-BP1/eIF4E Signalling Pathway as a Source of Cancer Drug Targets.Curr Med Chem. 2022;29(20):3501-3529. doi: 10.2174/0929867329666220224112042. Curr Med Chem. 2022. PMID: 35209811 Review.
Cited by
-
Mitotic CDK1 and 4E-BP1 II: A single phosphomimetic mutation in 4E-BP1 induces glucose intolerance in mice.PLoS One. 2023 Mar 10;18(3):e0282914. doi: 10.1371/journal.pone.0282914. eCollection 2023. PLoS One. 2023. PMID: 36897840 Free PMC article.
-
Deep computational analysis details dysregulation of eukaryotic translation initiation complex eIF4F in human cancers.Cell Syst. 2021 Sep 22;12(9):907-923.e6. doi: 10.1016/j.cels.2021.07.002. Epub 2021 Aug 5. Cell Syst. 2021. PMID: 34358439 Free PMC article.
-
Mitotic CDK1 and 4E-BP1 I: Loss of 4E-BP1 serine 82 phosphorylation promotes proliferative polycystic disease and lymphoma in aged or sublethally irradiated mice.PLoS One. 2023 May 5;18(5):e0282722. doi: 10.1371/journal.pone.0282722. eCollection 2023. PLoS One. 2023. PMID: 37145994 Free PMC article.
-
UV damage induces G3BP1-dependent stress granule formation that is not driven by mTOR inhibition-mediated translation arrest.J Cell Sci. 2020 Oct 28;133(20):jcs248310. doi: 10.1242/jcs.248310. J Cell Sci. 2020. PMID: 32989041 Free PMC article.
-
Inhibition of WNK Kinases in NK Cells Disrupts Cellular Osmoregulation and Control of Tumor Metastasis.J Innate Immun. 2024;16(1):451-469. doi: 10.1159/000540744. Epub 2024 Sep 12. J Innate Immun. 2024. PMID: 39265537 Free PMC article.
References
-
- Grifo J. A., Tahara S. M., Morgan M. A., Shatkin A. J., and Merrick W. C. (1983) New initiation factor activity required for globin mRNA translation. J. Biol. Chem. 258, 5804–5810 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
