

A New Suite of Allelic-Exchange Vectors for the Scarless Modification of Proteobacterial Genomes
- PMID: 31201277
- PMCID: PMC6677854
- DOI: 10.1128/AEM.00990-19
A New Suite of Allelic-Exchange Vectors for the Scarless Modification of Proteobacterial Genomes
Abstract
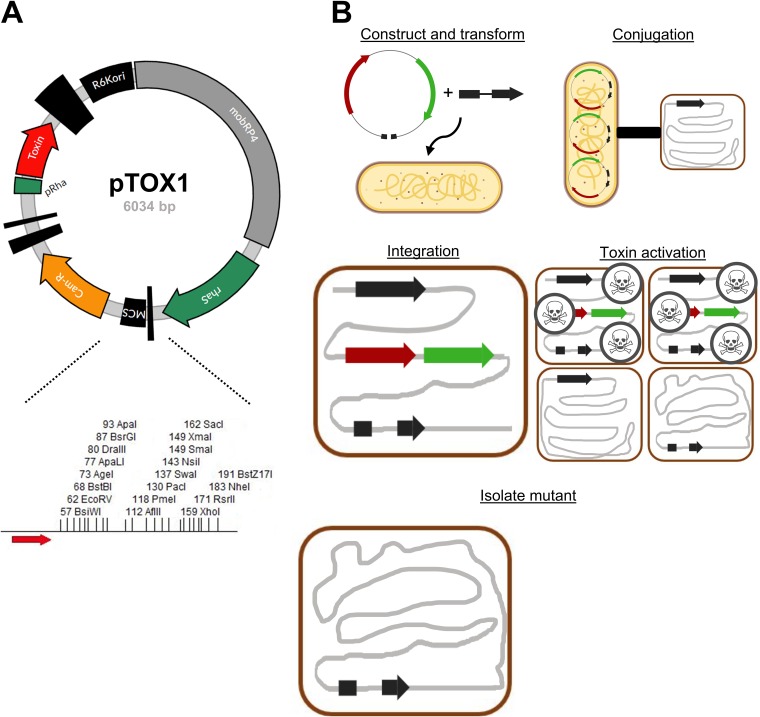
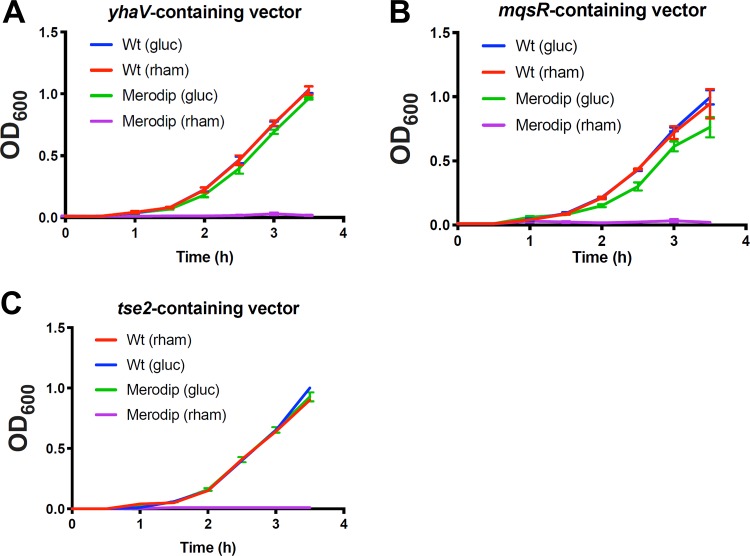
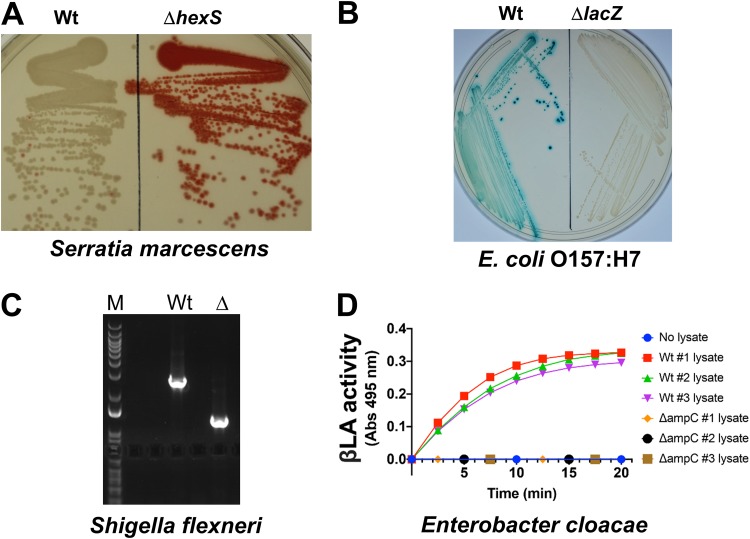
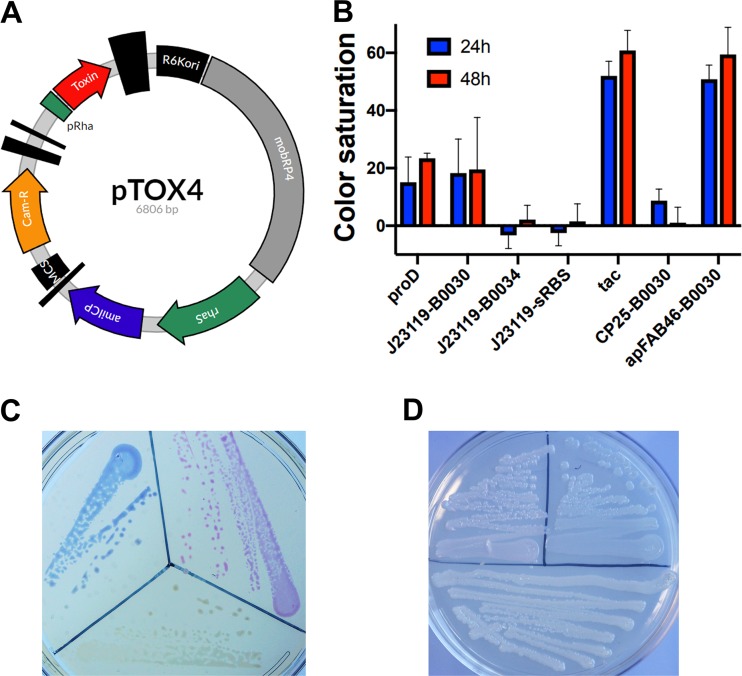
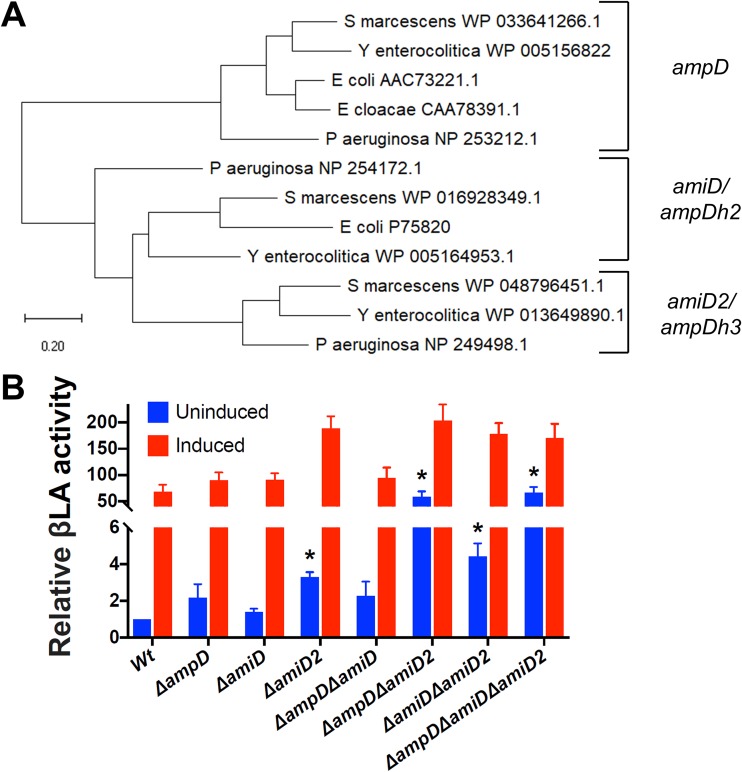
Despite the advent of new techniques for genetic engineering of bacteria, allelic exchange through homologous recombination remains an important tool for genetic analysis. Currently, sacB-based vector systems are often used for allelic exchange, but counterselection escape, which prevents isolation of cells with the desired mutation, occasionally limits their utility. To circumvent this, we engineered a series of "pTOX" allelic-exchange vectors. Each plasmid encodes one of a set of inducible toxins, chosen for their potential utility in a wide range of medically important proteobacteria. A codon-optimized rhaS transcriptional activator with a strong synthetic ribosome-binding site enables tight toxin induction even in organisms lacking an endogenous rhamnose regulon. Expression of the gene encoding blue AmilCP or magenta TsPurple nonfluorescent chromoprotein facilitates monitoring of successful single- and double-crossover events using these vectors. The versatility of these vectors was demonstrated by deleting genes in Serratia marcescens, Escherichia coli O157:H7, Enterobacter cloacae, and Shigella flexneri Finally, pTOX was used to characterize the impact of disruption of all combinations of the 3 paralogous S. marcescens peptidoglycan amidohydrolases on chromosomal ampC β-lactamase activity and the corresponding β-lactam antibiotic resistance. Mutation of multiple amidohydrolases was necessary for high-level ampC derepression and β-lactam resistance. These data suggest why β-lactam resistance may emerge during treatment less frequently in S. marcescens than in other AmpC-producing pathogens, like E. cloacae Collectively, our findings suggest that the pTOX vectors should be broadly useful for genetic engineering of Gram-negative bacteria.IMPORTANCE Targeted modification of bacterial genomes is critical for genetic analysis of microorganisms. Allelic exchange is a technique that relies on homologous recombination to replace native loci with engineered sequences. However, current allelic-exchange vectors often enable only weak selection for successful homologous recombination. We developed a suite of new allelic-exchange vectors, pTOX, which were validated in several medically important proteobacteria. They encode visible nonfluorescent chromoproteins that enable easy identification of colonies bearing integrated vectors and permit stringent selection for the second step of homologous recombination. We demonstrate the utility of these vectors by using them to investigate the effect of inactivation of Serratia marcescens peptidoglycan amidohydrolases on β-lactam antibiotic resistance.
Keywords: AmilCP gene; Serratia marcescens; allelic exchange; ampC; ampD; antibiotic resistance; beta-lactamase; sacB; toxin-antitoxin; type VI toxin.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Complex Regulation Pathways of AmpC-Mediated β-Lactam Resistance in Enterobacter cloacae Complex.Antimicrob Agents Chemother. 2015 Dec;59(12):7753-61. doi: 10.1128/AAC.01729-15. Epub 2015 Oct 5. Antimicrob Agents Chemother. 2015. PMID: 26438498 Free PMC article.
-
Divergent genetic landscapes drive lower levels of AmpC induction and stable de-repression in Serratia marcescens compared to Enterobacter cloacae.Antimicrob Agents Chemother. 2024 Jan 10;68(1):e0119323. doi: 10.1128/aac.01193-23. Epub 2023 Dec 12. Antimicrob Agents Chemother. 2024. PMID: 38084952 Free PMC article.
-
New cloning vectors to facilitate quick allelic exchange in gram-negative bacteria.Biotechniques. 2021 Feb;70(2):116-119. doi: 10.2144/btn-2020-0135. Epub 2021 Jan 25. Biotechniques. 2021. PMID: 33492170
-
A Primer on AmpC β-Lactamases: Necessary Knowledge for an Increasingly Multidrug-resistant World.Clin Infect Dis. 2019 Sep 27;69(8):1446-1455. doi: 10.1093/cid/ciz173. Clin Infect Dis. 2019. PMID: 30838380 Free PMC article. Review.
-
AmpC beta-lactamases.Clin Microbiol Rev. 2009 Jan;22(1):161-82, Table of Contents. doi: 10.1128/CMR.00036-08. Clin Microbiol Rev. 2009. PMID: 19136439 Free PMC article. Review.
Cited by
-
Disrupting Central Carbon Metabolism Increases β-Lactam Antibiotic Susceptibility in Vibrio cholerae.J Bacteriol. 2023 Mar 21;205(3):e0047622. doi: 10.1128/jb.00476-22. Epub 2023 Feb 22. J Bacteriol. 2023. PMID: 36840595 Free PMC article.
-
Versatile allelic replacement and self-excising integrative vectors for plasmid genome mutation and complementation.Microbiol Spectr. 2024 Jan 11;12(1):e0338723. doi: 10.1128/spectrum.03387-23. Epub 2023 Nov 22. Microbiol Spectr. 2024. PMID: 37991378 Free PMC article.
-
Multiple resistance factors collectively promote inoculum-dependent dynamic survival during antimicrobial peptide exposure in Enterobacter cloacae.bioRxiv [Preprint]. 2024 Mar 3:2024.03.03.583169. doi: 10.1101/2024.03.03.583169. bioRxiv. 2024. Update in: PLoS Pathog. 2024 Aug 26;20(8):e1012488. doi: 10.1371/journal.ppat.1012488. PMID: 38463991 Free PMC article. Updated. Preprint.
-
Mobilizable shuttle vectors with fluorescent markers functional across different species of bacteria.Appl Environ Microbiol. 2025 Jun 18;91(6):e0004525. doi: 10.1128/aem.00045-25. Epub 2025 May 12. Appl Environ Microbiol. 2025. PMID: 40353662 Free PMC article.
-
Genetic Manipulation of Wild Human Gut Bacteroides.J Bacteriol. 2020 Jan 15;202(3):e00544-19. doi: 10.1128/JB.00544-19. Print 2020 Jan 15. J Bacteriol. 2020. PMID: 31712278 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
