

Analysis of Host Responses to Hepatitis B and Delta Viral Infections in a Micro-scalable Hepatic Co-culture System
- PMID: 31206195
- PMCID: PMC6917996
- DOI: 10.1002/hep.30815
Analysis of Host Responses to Hepatitis B and Delta Viral Infections in a Micro-scalable Hepatic Co-culture System
Abstract
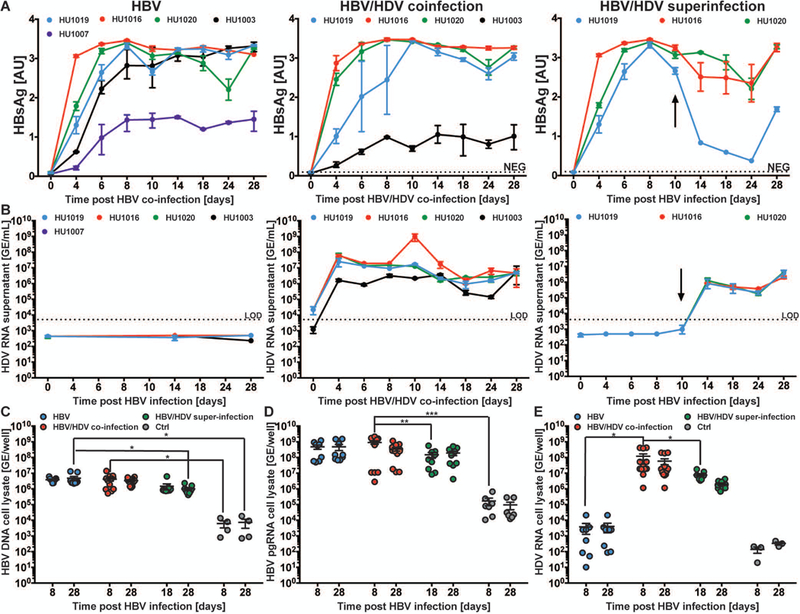
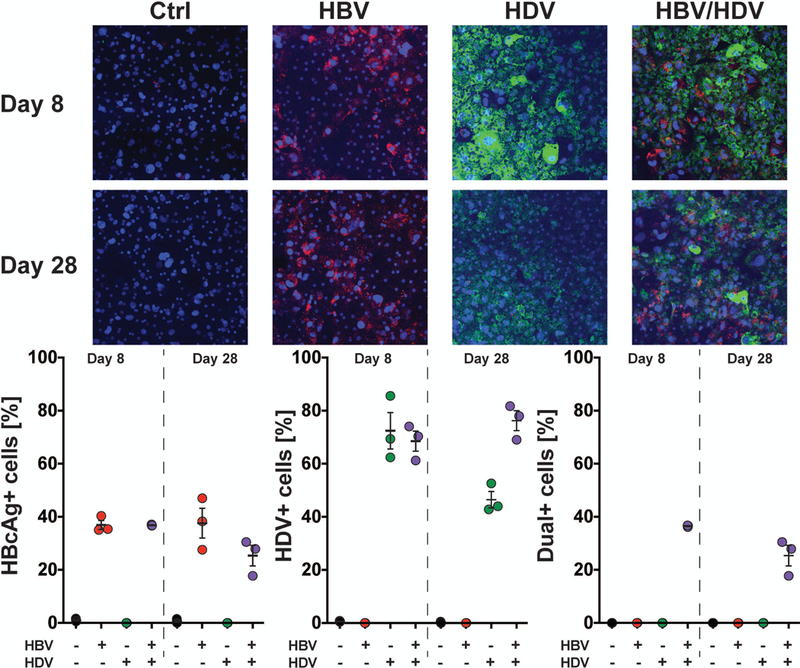
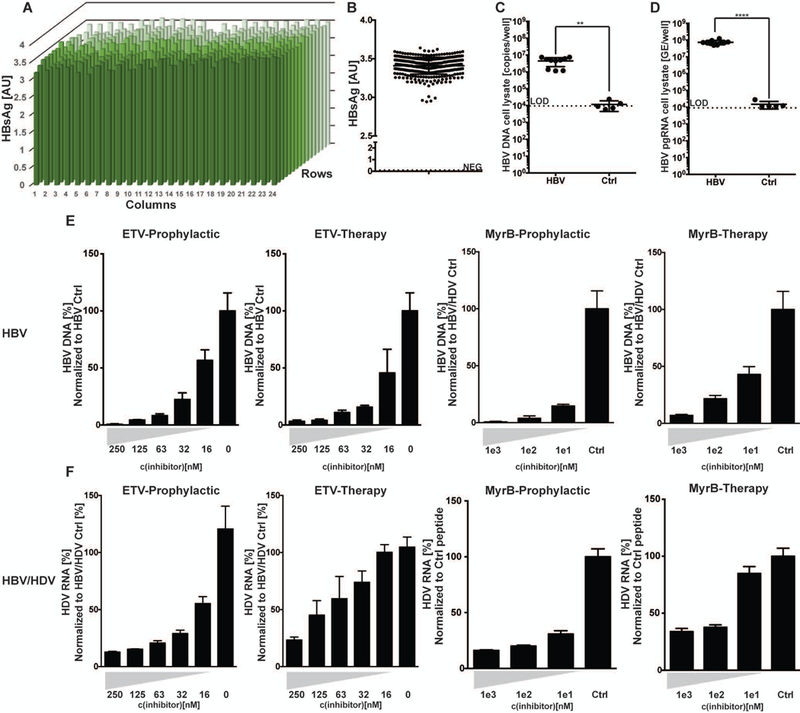
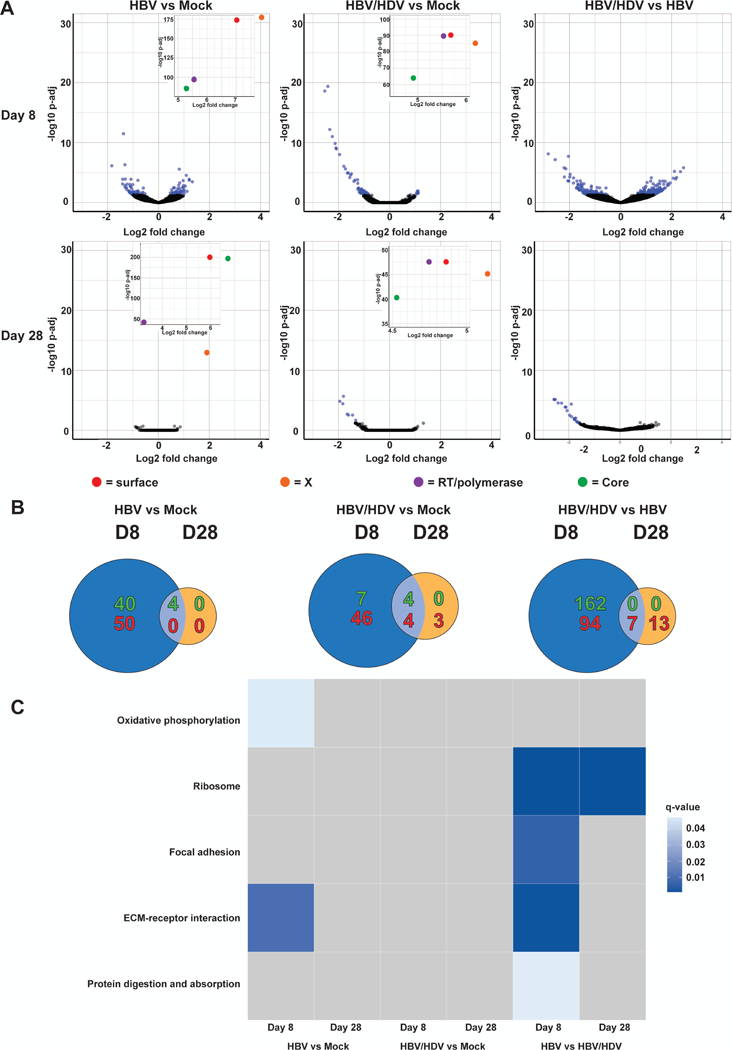
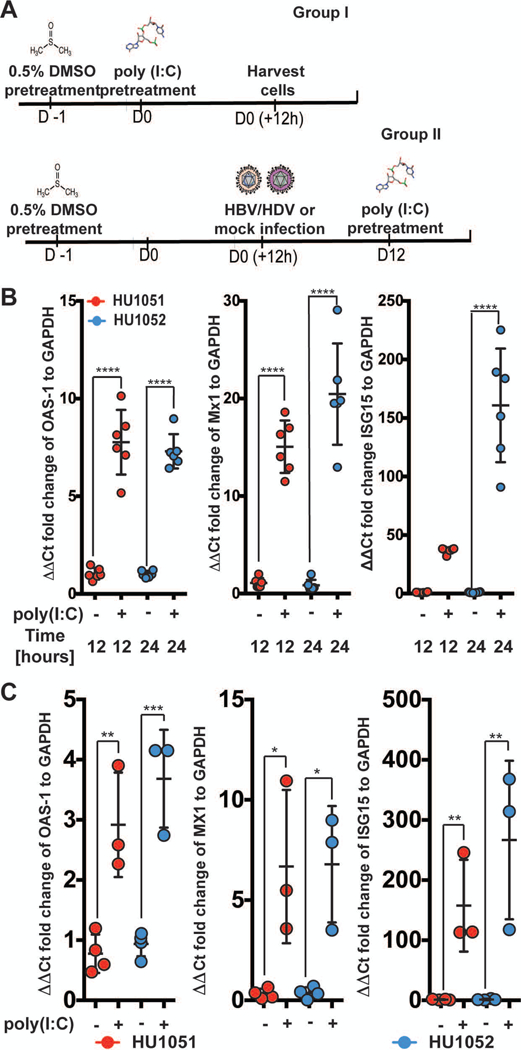
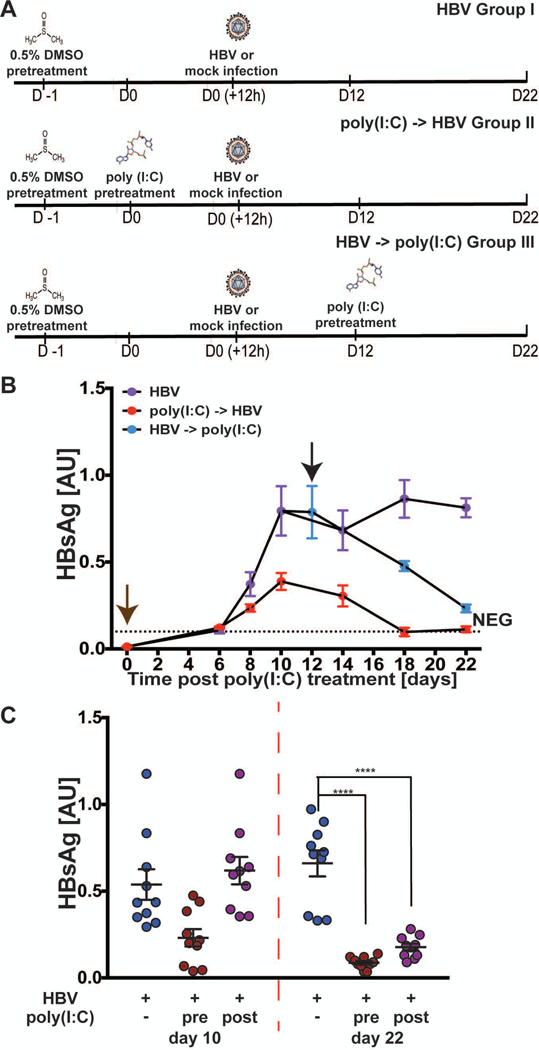
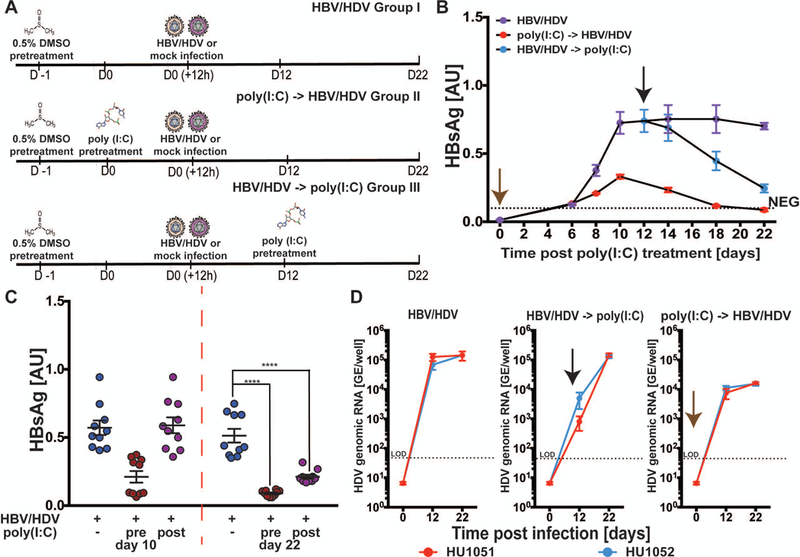
Hepatitis B virus (HBV) remains a major global health problem with 257 million chronically infected individuals worldwide, of whom approximately 20 million are co-infected with hepatitis delta virus (HDV). Progress toward a better understanding of the complex interplay between these two viruses and the development of novel therapies have been hampered by the scarcity of suitable cell culture models that mimic the natural environment of the liver. Here, we established HBV and HBV/HDV co-infections and super-infections in self-assembling co-cultured primary human hepatocytes (SACC-PHHs) for up to 28 days in a 384-well format and highlight the suitability of this platform for high-throughput drug testing. We performed RNA sequencing at days 8 and 28 on SACC-PHHs, either HBV mono-infected or HBV/HDV co-infected. Our transcriptomic analysis demonstrates that hepatocytes in SACC-PHHs maintain a mature hepatic phenotype over time, regardless of infection condition. We confirm that HBV is a stealth virus, as it does not induce a strong innate immune response; rather, oxidative phosphorylation and extracellular matrix-receptor interactions are dysregulated to create an environment that promotes persistence. Notably, HDV co-infection also did not lead to statistically significant transcriptional changes across multiple donors and replicates. The lack of innate immune activation is not due to SACC-PHHs being impaired in their ability to induce interferon stimulated genes (ISGs). Rather, polyinosinic:polycytidylic acid exposure activates ISGs, and this stimulation significantly inhibits HBV infection, yet only minimally affects the ability of HDV to infect and persist. Conclusion: These data demonstrate that the SACC-PHH system is a versatile platform for studying HBV/HDV co-infections and holds promise for performing chemical library screens and improving our understanding of the host response to such infections.
© 2019 by the American Association for the Study of Liver Diseases.
Conflict of interest statement
Conflicts of Interest.
The following authors declare competing financial interests. R.J., Am.P., C.C., A.S. are employees of the Hurel Corporation of which E.N. is also a stockholder. A.P. is a member of the Scientific Advisory Board of the Hurel Corporation. B.Y.W., J.M.G., Y.B., R.E.S., G.E.L., L.P., and R.L. do not have any conflicts of interest.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
