

Regulation of ectopic heterochromatin-mediated epigenetic diversification by the JmjC family protein Epe1
- PMID: 31206516
- PMCID: PMC6576747
- DOI: 10.1371/journal.pgen.1008129
Regulation of ectopic heterochromatin-mediated epigenetic diversification by the JmjC family protein Epe1
Abstract
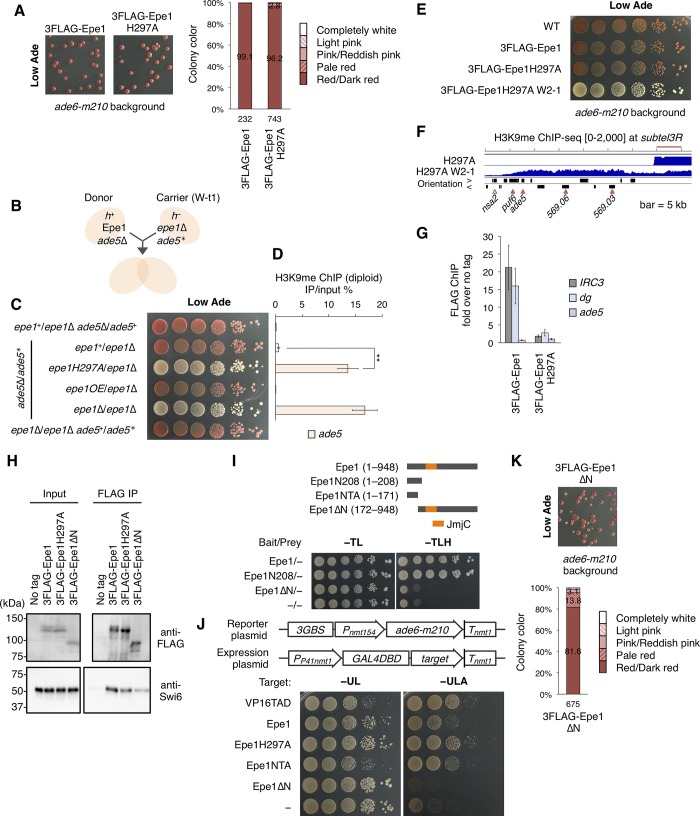
H3K9 methylation (H3K9me) is a conserved marker of heterochromatin, a transcriptionally silent chromatin structure. Knowledge of the mechanisms for regulating heterochromatin distribution is limited. The fission yeast JmjC domain-containing protein Epe1 localizes to heterochromatin mainly through its interaction with Swi6, a homologue of heterochromatin protein 1 (HP1), and directs JmjC-mediated H3K9me demethylation in vivo. Here, we found that loss of epe1 (epe1Δ) induced a red-white variegated phenotype in a red-pigment accumulation background that generated uniform red colonies. Analysis of isolated red and white colonies revealed that silencing of genes involved in pigment accumulation by stochastic ectopic heterochromatin formation led to white colony formation. In addition, genome-wide analysis of red- and white-isolated clones revealed that epe1Δ resulted in a heterogeneous heterochromatin distribution among clones. We found that Epe1 had an N-terminal domain distinct from its JmjC domain, which activated transcription in both fission and budding yeasts. The N-terminal transcriptional activation (NTA) domain was involved in suppression of ectopic heterochromatin-mediated red-white variegation. We introduced a single copy of Epe1 into epe1Δ clones harboring ectopic heterochromatin, and found that Epe1 could reduce H3K9me from ectopic heterochromatin but some of the heterochromatin persisted. This persistence was due to a latent H3K9me source embedded in ectopic heterochromatin. Epe1H297A, a canonical JmjC mutant, suppressed red-white variegation, but entirely failed to remove already-established ectopic heterochromatin, suggesting that Epe1 prevented stochastic de novo deposition of ectopic H3K9me in an NTA-dependent but JmjC-independent manner, while its JmjC domain mediated removal of H3K9me from established ectopic heterochromatin. Our results suggest that Epe1 not only limits the distribution of heterochromatin but also controls the balance between suppression and retention of heterochromatin-mediated epigenetic diversification.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
An H3K9 methylation-dependent protein interaction regulates the non-enzymatic functions of a putative histone demethylase.Elife. 2020 Mar 20;9:e53155. doi: 10.7554/eLife.53155. Elife. 2020. PMID: 32195666 Free PMC article.
-
Interaction of Epe1 with the heterochromatin assembly pathway in Schizosaccharomyces pombe.Genetics. 2007 Apr;175(4):1549-60. doi: 10.1534/genetics.106.068684. Genetics. 2007. PMID: 17449867 Free PMC article.
-
The JmjC domain protein Epe1 prevents unregulated assembly and disassembly of heterochromatin.EMBO J. 2007 Nov 14;26(22):4670-82. doi: 10.1038/sj.emboj.7601892. Epub 2007 Oct 18. EMBO J. 2007. PMID: 17948055 Free PMC article.
-
Unprogrammed epigenetic variation mediated by stochastic formation of ectopic heterochromatin.Curr Genet. 2020 Apr;66(2):319-325. doi: 10.1007/s00294-019-01031-4. Epub 2019 Oct 9. Curr Genet. 2020. PMID: 31598751 Review.
-
Destabilizing heterochromatin: Does Swi6/HP1 make the choice?Mol Cell. 2006 Jun 23;22(6):709-710. doi: 10.1016/j.molcel.2006.06.004. Mol Cell. 2006. PMID: 16793539 Review.
Cited by
-
Opposing Roles of FACT for Euchromatin and Heterochromatin in Yeast.Biomolecules. 2023 Feb 16;13(2):377. doi: 10.3390/biom13020377. Biomolecules. 2023. PMID: 36830746 Free PMC article. Review.
-
An H3K9 methylation-dependent protein interaction regulates the non-enzymatic functions of a putative histone demethylase.Elife. 2020 Mar 20;9:e53155. doi: 10.7554/eLife.53155. Elife. 2020. PMID: 32195666 Free PMC article.
-
Epigenetic gene silencing by heterochromatin primes fungal resistance.Nature. 2020 Sep;585(7825):453-458. doi: 10.1038/s41586-020-2706-x. Epub 2020 Sep 9. Nature. 2020. PMID: 32908306 Free PMC article.
-
A conserved RNA degradation complex required for spreading and epigenetic inheritance of heterochromatin.Elife. 2020 Jun 3;9:e54341. doi: 10.7554/eLife.54341. Elife. 2020. PMID: 32491985 Free PMC article.
-
Mapping the dynamics of epigenetic adaptation in S. pombe during heterochromatin misregulation.Dev Cell. 2024 Aug 19;59(16):2222-2238.e4. doi: 10.1016/j.devcel.2024.07.006. Epub 2024 Aug 1. Dev Cell. 2024. PMID: 39094565 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
