

Perturbation of synapsins homeostasis through HIV-1 Tat-mediated suppression of BAG3 in primary neuronal cells
- PMID: 31209204
- PMCID: PMC6572798
- DOI: 10.1038/s41419-019-1702-2
Perturbation of synapsins homeostasis through HIV-1 Tat-mediated suppression of BAG3 in primary neuronal cells
Abstract
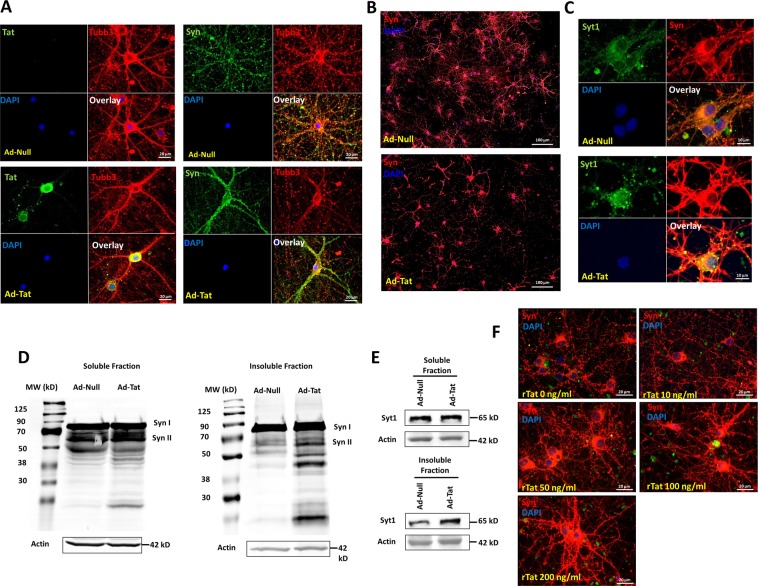
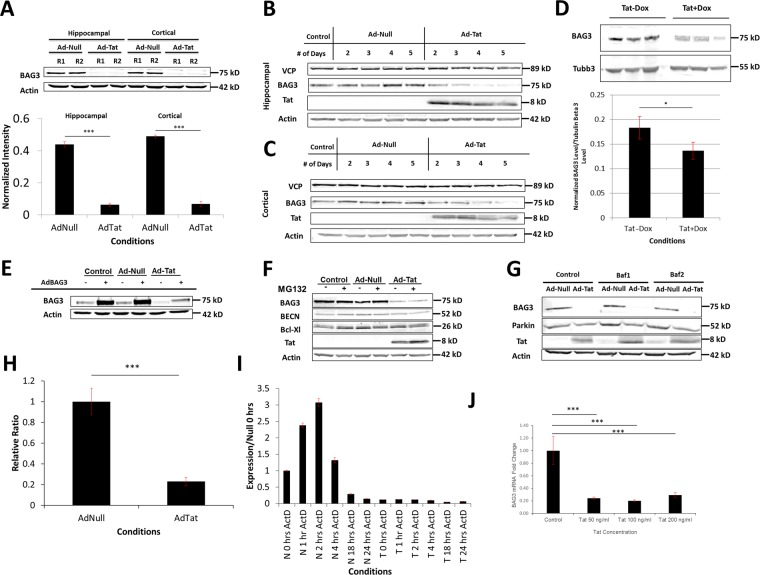
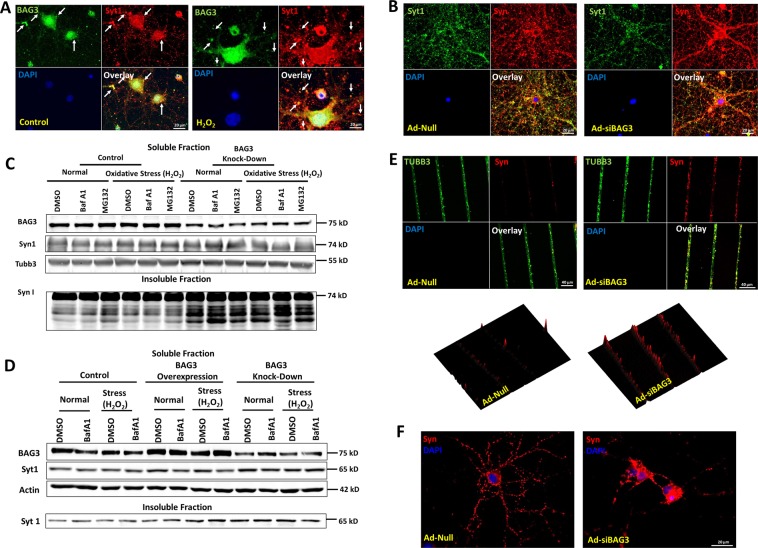
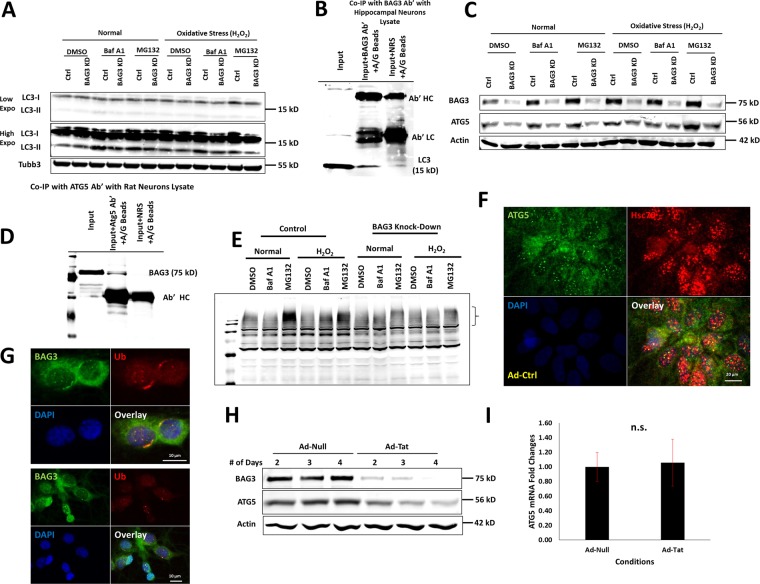
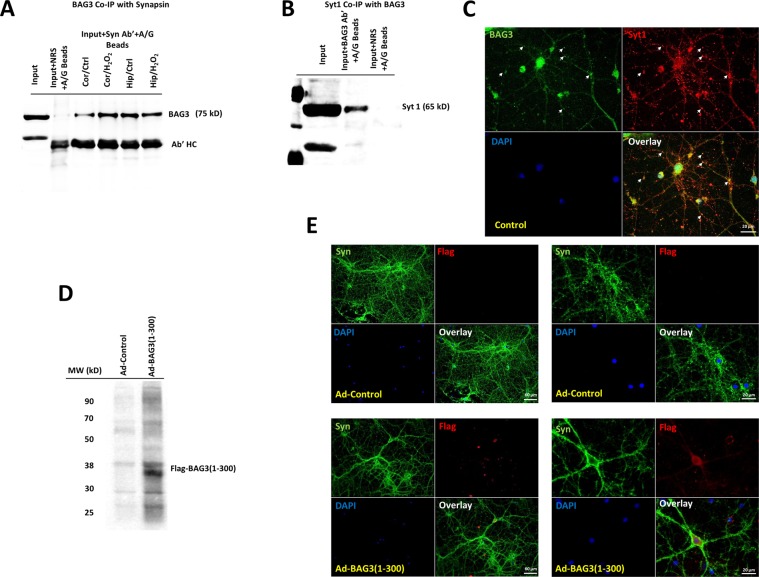
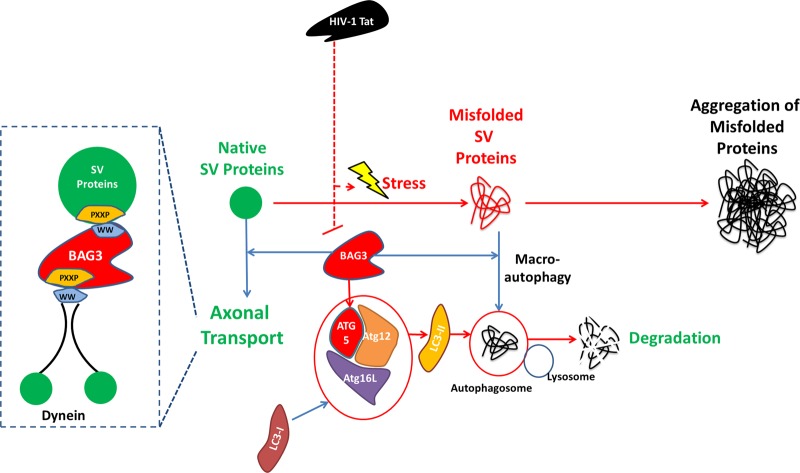
HIV-1 Tat is known to be released by HIV infected non-neuronal cells in the brain, and after entering neurons, compromises brain homeostasis by impairing pro-survival pathways, thus contributing to the development of HIV-associated CNS disorders commonly observed in individuals living with HIV. Here, we demonstrate that synapsins, phosphoproteins that are predominantly expressed in neuronal cells and play a vital role in modulating neurotransmitter release at the pre-synaptic terminal, and neuronal differentiation become targets for Tat through autophagy and protein quality control pathways. We demonstrate that the presence of Tat in neurons results in downregulation of BAG3, a co-chaperone for heat shock proteins (Hsp70/Hsc70) that is implicated in protein quality control (PQC) processes by eliminating mis-folded and damaged proteins, and selective macroautophagy. Our results show that treatment of cells with Tat or suppression of BAG3 expression by siRNA in neuronal cells disturbs subcellular distribution of synapsins and synaptotagmin 1 (Syt1) leading to their accumulation in the neuronal soma and along axons in a punctate pattern, rather than being properly distributed at axon-terminals. Further, our results revealed that synapsins partially lost their stability and their removal via lysosomal autophagy was noticeably impaired in cells with low levels of BAG3. The observed impairment of lysosomal autophagy, under this condition, is likely caused by cells losing their ability to process LC3-I to LC3-II, in part due to a decrease in the ATG5 levels upon BAG3 knockdown. These observations ascribe a new function for BAG3 in controlling synaptic communications and illuminate a new downstream target for Tat to elicit its pathogenic effect in impacting neuronal cell function and behavior.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
HIV-1 Tat increases BAG3 via NF-κB signaling to induce autophagy during HIV-associated neurocognitive disorder.Cell Cycle. 2018;17(13):1614-1623. doi: 10.1080/15384101.2018.1480219. Epub 2018 Aug 21. Cell Cycle. 2018. PMID: 29962275 Free PMC article.
-
Enhanced autophagic-lysosomal activity and increased BAG3-mediated selective macroautophagy as adaptive response of neuronal cells to chronic oxidative stress.Redox Biol. 2019 Jun;24:101181. doi: 10.1016/j.redox.2019.101181. Epub 2019 Apr 2. Redox Biol. 2019. PMID: 30959460 Free PMC article.
-
BAG3 and SYNPO (synaptopodin) facilitate phospho-MAPT/Tau degradation via autophagy in neuronal processes.Autophagy. 2019 Jul;15(7):1199-1213. doi: 10.1080/15548627.2019.1580096. Epub 2019 Mar 1. Autophagy. 2019. PMID: 30744518 Free PMC article.
-
Breaking BAG: The Co-Chaperone BAG3 in Health and Disease.Trends Pharmacol Sci. 2016 Aug;37(8):672-688. doi: 10.1016/j.tips.2016.04.007. Epub 2016 May 6. Trends Pharmacol Sci. 2016. PMID: 27162137 Review.
-
Unraveling the mystery: How bad is BAG3 in hematological malignancies?Biochim Biophys Acta Rev Cancer. 2022 Sep;1877(5):188781. doi: 10.1016/j.bbcan.2022.188781. Epub 2022 Aug 17. Biochim Biophys Acta Rev Cancer. 2022. PMID: 35985611 Review.
Cited by
-
Cross-talk between lipid homeostasis and endoplasmic reticulum stress in neurodegeneration: Insights for HIV-1 associated neurocognitive disorders (HAND).Neurochem Int. 2020 Dec;141:104880. doi: 10.1016/j.neuint.2020.104880. Epub 2020 Oct 14. Neurochem Int. 2020. PMID: 33065212 Free PMC article. Review.
-
Chaperone-Mediated Autophagy in Brain Injury: A Double-Edged Sword with Therapeutic Potentials.Mol Neurobiol. 2024 Dec;61(12):10671-10683. doi: 10.1007/s12035-024-04230-4. Epub 2024 May 22. Mol Neurobiol. 2024. PMID: 38775879 Review.
-
Systems-Level Interactome Mapping Reveals Actionable Protein Network Dysregulation Across the Alzheimer's Disease Spectrum.Res Sq [Preprint]. 2025 Feb 12:rs.3.rs-5930673. doi: 10.21203/rs.3.rs-5930673/v1. Res Sq. 2025. PMID: 39989971 Free PMC article. Preprint.
-
Acute Administration of HIV-1 Tat Protein Drives Glutamatergic Alterations in a Rodent Model of HIV-Associated Neurocognitive Disorders.Mol Neurobiol. 2024 Oct;61(10):8467-8480. doi: 10.1007/s12035-024-04113-8. Epub 2024 Mar 22. Mol Neurobiol. 2024. PMID: 38514527 Free PMC article.
-
Neuromodulation of BAG co-chaperones by HIV-1 viral proteins and H2O2: implications for HIV-associated neurological disorders.Cell Death Discov. 2021 Mar 26;7(1):60. doi: 10.1038/s41420-021-00424-0. Cell Death Discov. 2021. PMID: 33771978 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
