

Mediator complex interaction partners organize the transcriptional network that defines neural stem cells
- PMID: 31209209
- PMCID: PMC6573065
- DOI: 10.1038/s41467-019-10502-8
Mediator complex interaction partners organize the transcriptional network that defines neural stem cells
Erratum in
-
Publisher Correction: Mediator complex interaction partners organize the transcriptional network that defines neural stem cells.Nat Commun. 2019 Jul 22;10(1):3318. doi: 10.1038/s41467-019-11254-1. Nat Commun. 2019. PMID: 31332183 Free PMC article.
Abstract
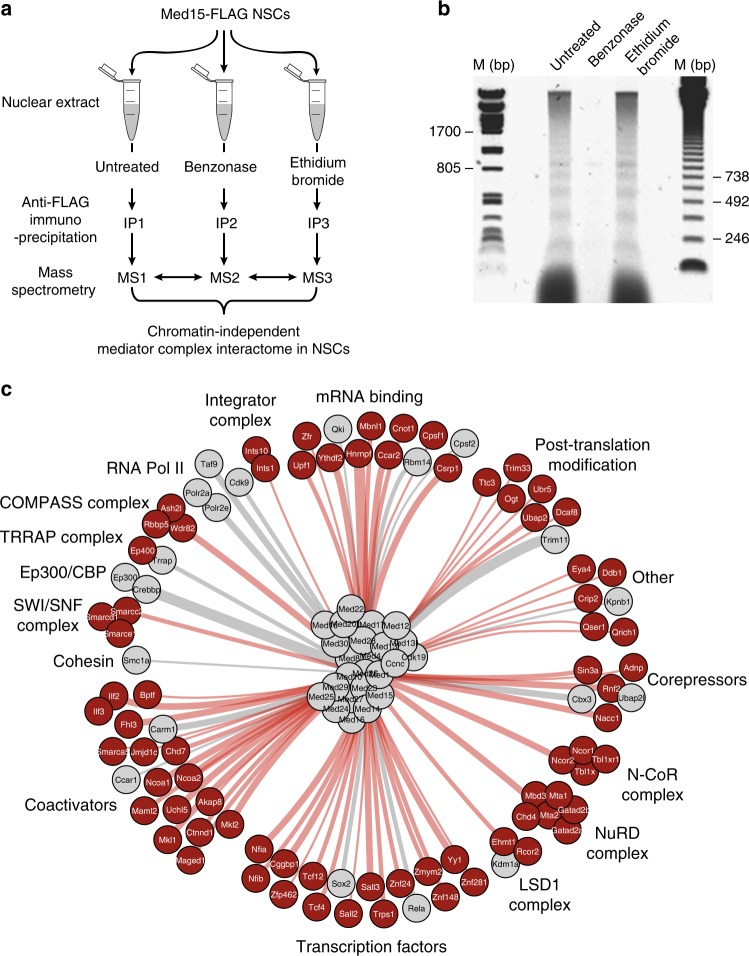
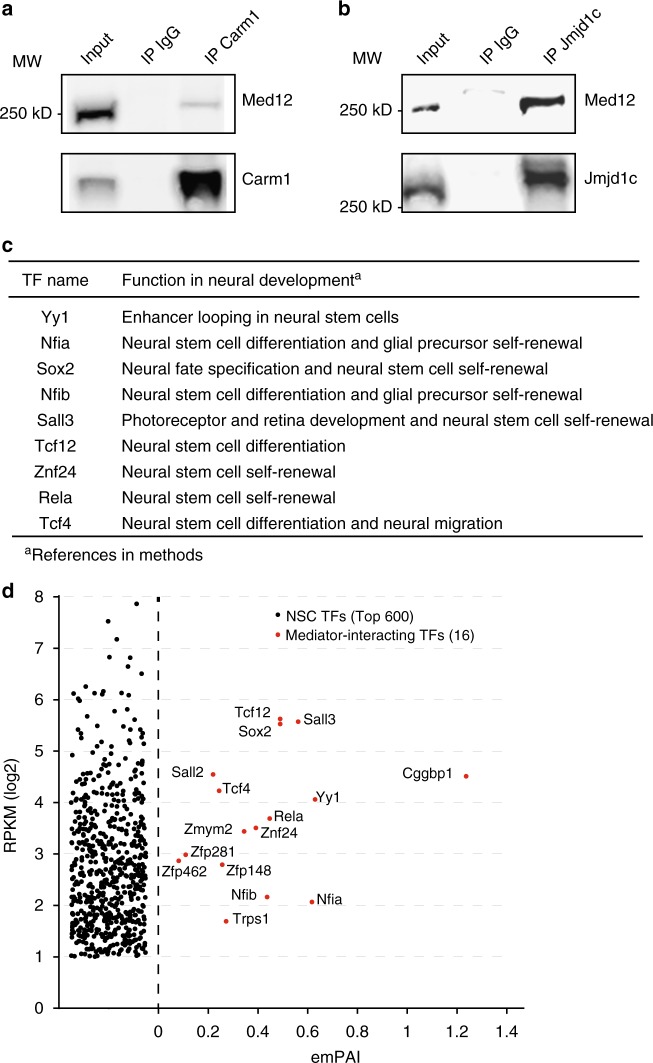
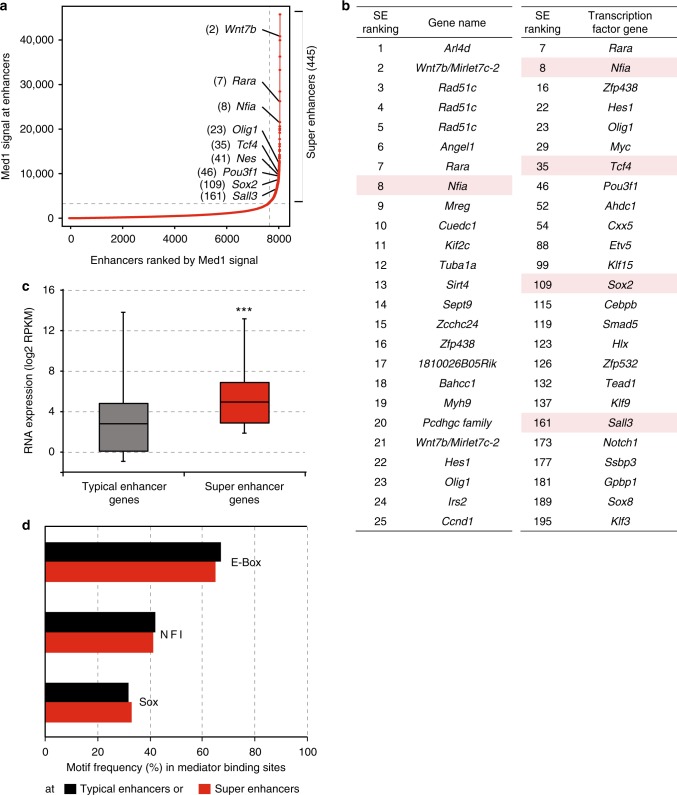
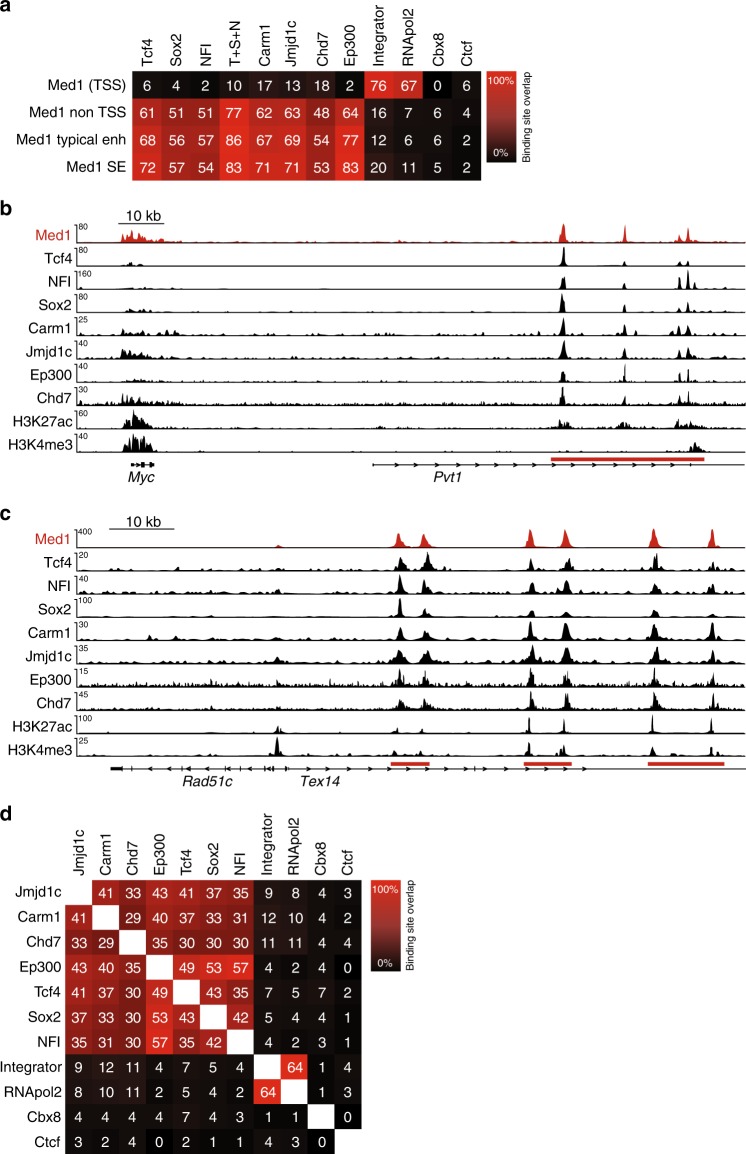
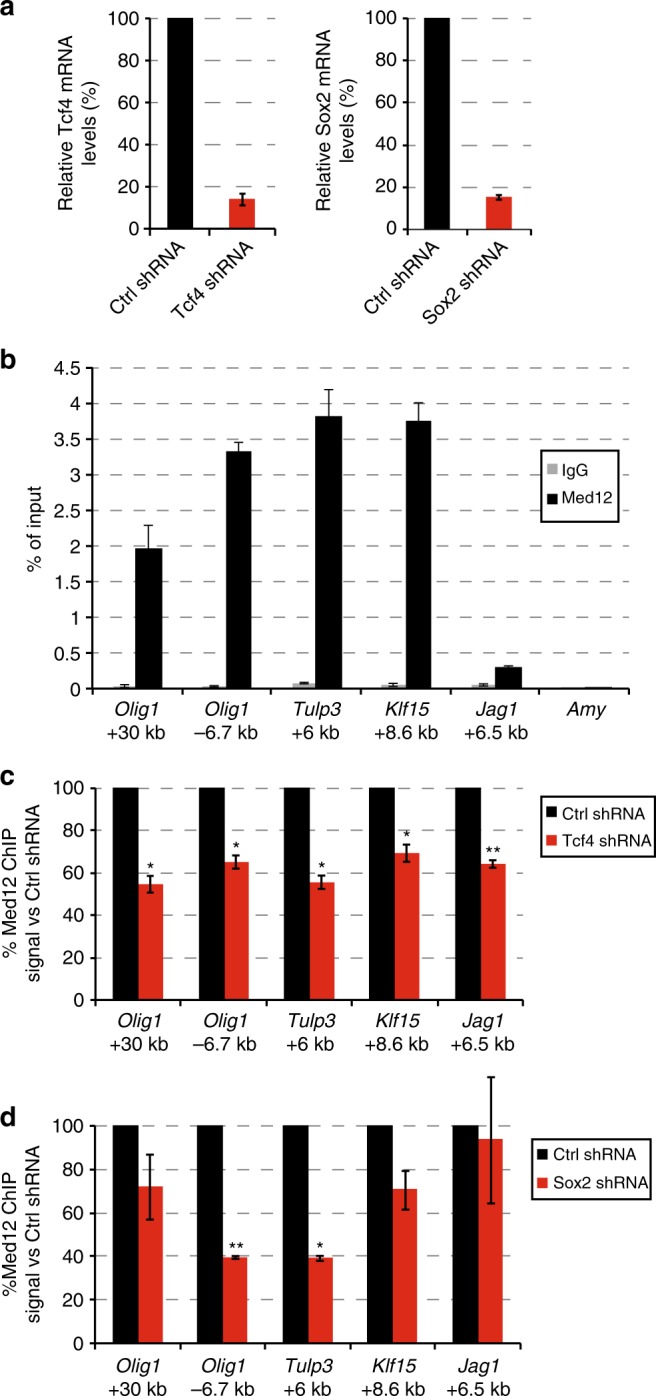
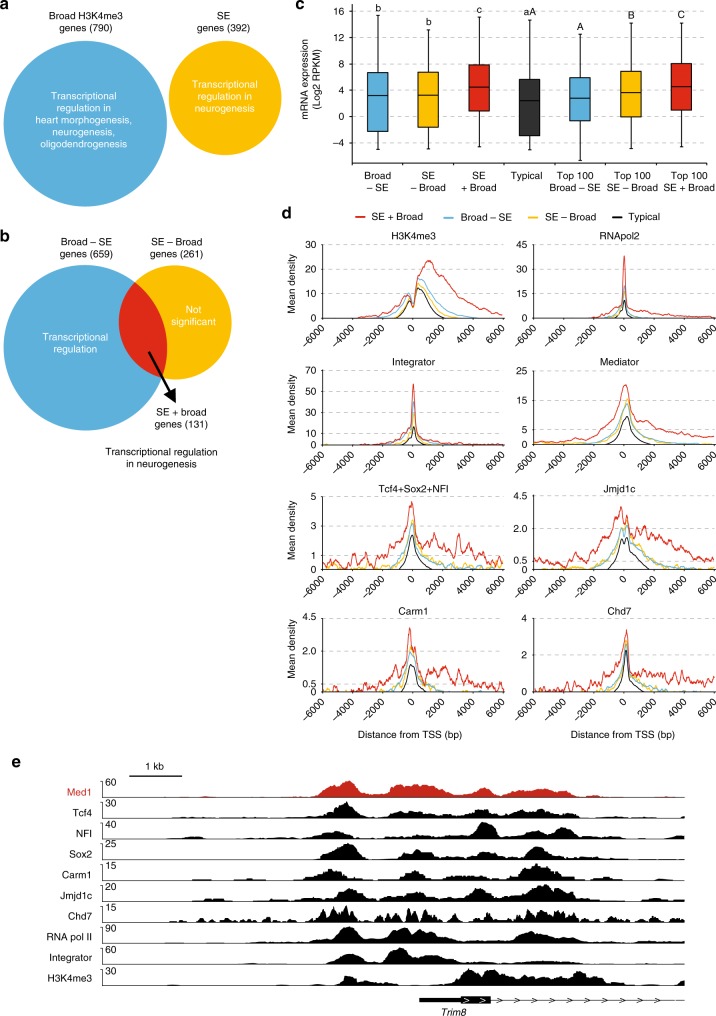
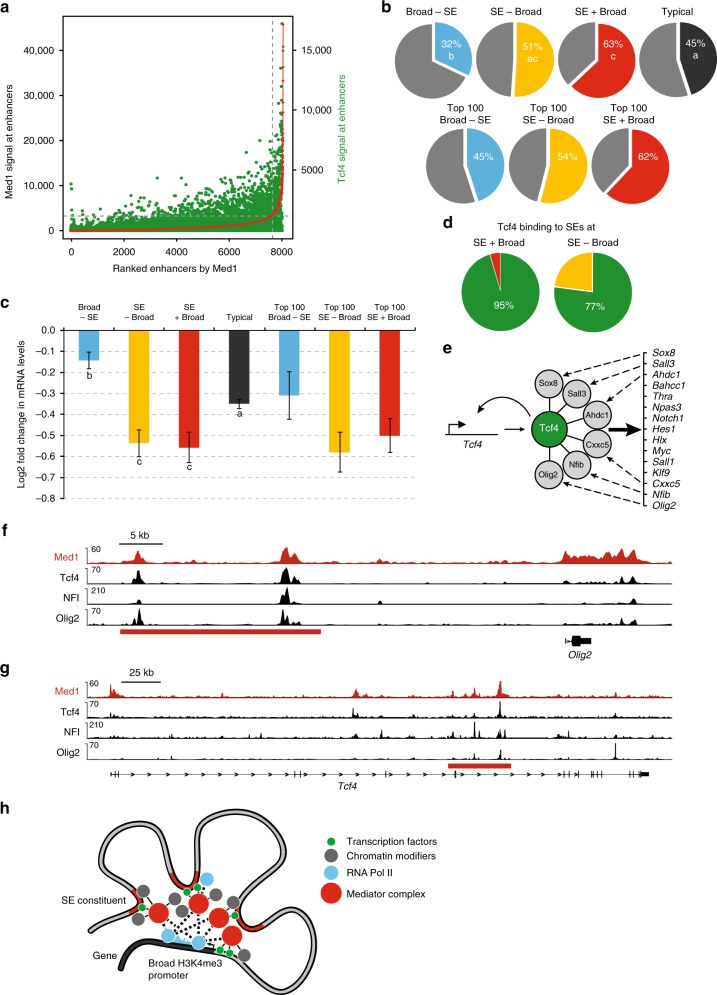
The Mediator complex regulates transcription by connecting enhancers to promoters. High Mediator binding density defines super enhancers, which regulate cell-identity genes and oncogenes. Protein interactions of Mediator may explain its role in these processes but have not been identified comprehensively. Here, we purify Mediator from neural stem cells (NSCs) and identify 75 protein-protein interaction partners. We identify super enhancers in NSCs and show that Mediator-interacting chromatin modifiers colocalize with Mediator at enhancers and super enhancers. Transcription factor families with high affinity for Mediator dominate enhancers and super enhancers and can explain genome-wide Mediator localization. We identify E-box transcription factor Tcf4 as a key regulator of NSCs. Tcf4 interacts with Mediator, colocalizes with Mediator at super enhancers and regulates neurogenic transcription factor genes with super enhancers and broad H3K4me3 domains. Our data suggest that high binding-affinity for Mediator is an important organizing feature in the transcriptional network that determines NSC identity.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Master transcription factors and mediator establish super-enhancers at key cell identity genes.Cell. 2013 Apr 11;153(2):307-19. doi: 10.1016/j.cell.2013.03.035. Cell. 2013. PMID: 23582322 Free PMC article.
-
Chromatin interaction networks revealed unique connectivity patterns of broad H3K4me3 domains and super enhancers in 3D chromatin.Sci Rep. 2017 Oct 31;7(1):14466. doi: 10.1038/s41598-017-14389-7. Sci Rep. 2017. PMID: 29089515 Free PMC article.
-
Enhancer RNAs: a missing regulatory layer in gene transcription.Sci China Life Sci. 2019 Jul;62(7):905-912. doi: 10.1007/s11427-017-9370-9. Epub 2018 Dec 26. Sci China Life Sci. 2019. PMID: 30593613 Review.
-
Super-Enhancers and Broad H3K4me3 Domains Form Complex Gene Regulatory Circuits Involving Chromatin Interactions.Sci Rep. 2017 May 19;7(1):2186. doi: 10.1038/s41598-017-02257-3. Sci Rep. 2017. PMID: 28526829 Free PMC article.
-
Broad H3K4me3 domains: Maintaining cellular identity and their implication in super-enhancer hijacking.Bioessays. 2023 Oct;45(10):e2200239. doi: 10.1002/bies.202200239. Epub 2023 Jun 23. Bioessays. 2023. PMID: 37350339 Review.
Cited by
-
Rapid profiling of transcription factor-cofactor interaction networks reveals principles of epigenetic regulation.Nucleic Acids Res. 2024 Sep 23;52(17):10276-10296. doi: 10.1093/nar/gkae706. Nucleic Acids Res. 2024. PMID: 39166482 Free PMC article.
-
The dynamic broad epigenetic (H3K4me3, H3K27ac) domain as a mark of essential genes.Clin Epigenetics. 2021 Jul 8;13(1):138. doi: 10.1186/s13148-021-01126-1. Clin Epigenetics. 2021. PMID: 34238359 Free PMC article. Review.
-
What do Transcription Factors Interact With?J Mol Biol. 2021 Jul 9;433(14):166883. doi: 10.1016/j.jmb.2021.166883. Epub 2021 Feb 20. J Mol Biol. 2021. PMID: 33621520 Free PMC article. Review.
-
Suppression of p53 response by targeting p53-Mediator binding with a stapled peptide.Cell Rep. 2022 Apr 5;39(1):110630. doi: 10.1016/j.celrep.2022.110630. Cell Rep. 2022. PMID: 35385747 Free PMC article.
-
Incomplete erasure of histone marks during epigenetic reprogramming in medaka early development.Genome Res. 2023 Apr;33(4):572-586. doi: 10.1101/gr.277577.122. Epub 2023 Apr 28. Genome Res. 2023. PMID: 37117034 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
