

A temporal thymic selection switch and ligand binding kinetics constrain neonatal Foxp3+ Treg cell development
- PMID: 31209405
- PMCID: PMC7039339
- DOI: 10.1038/s41590-019-0414-1
A temporal thymic selection switch and ligand binding kinetics constrain neonatal Foxp3+ Treg cell development
Abstract
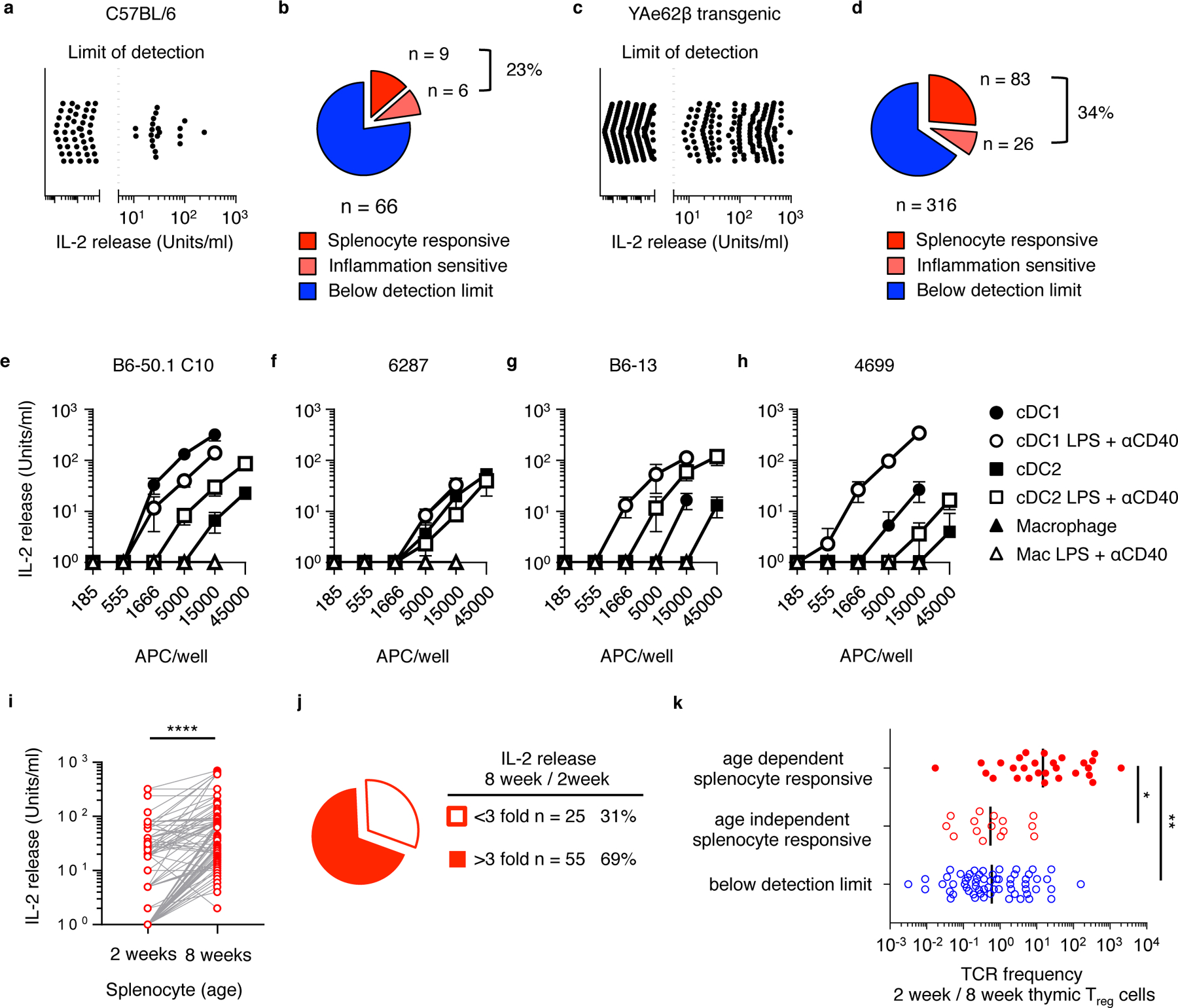
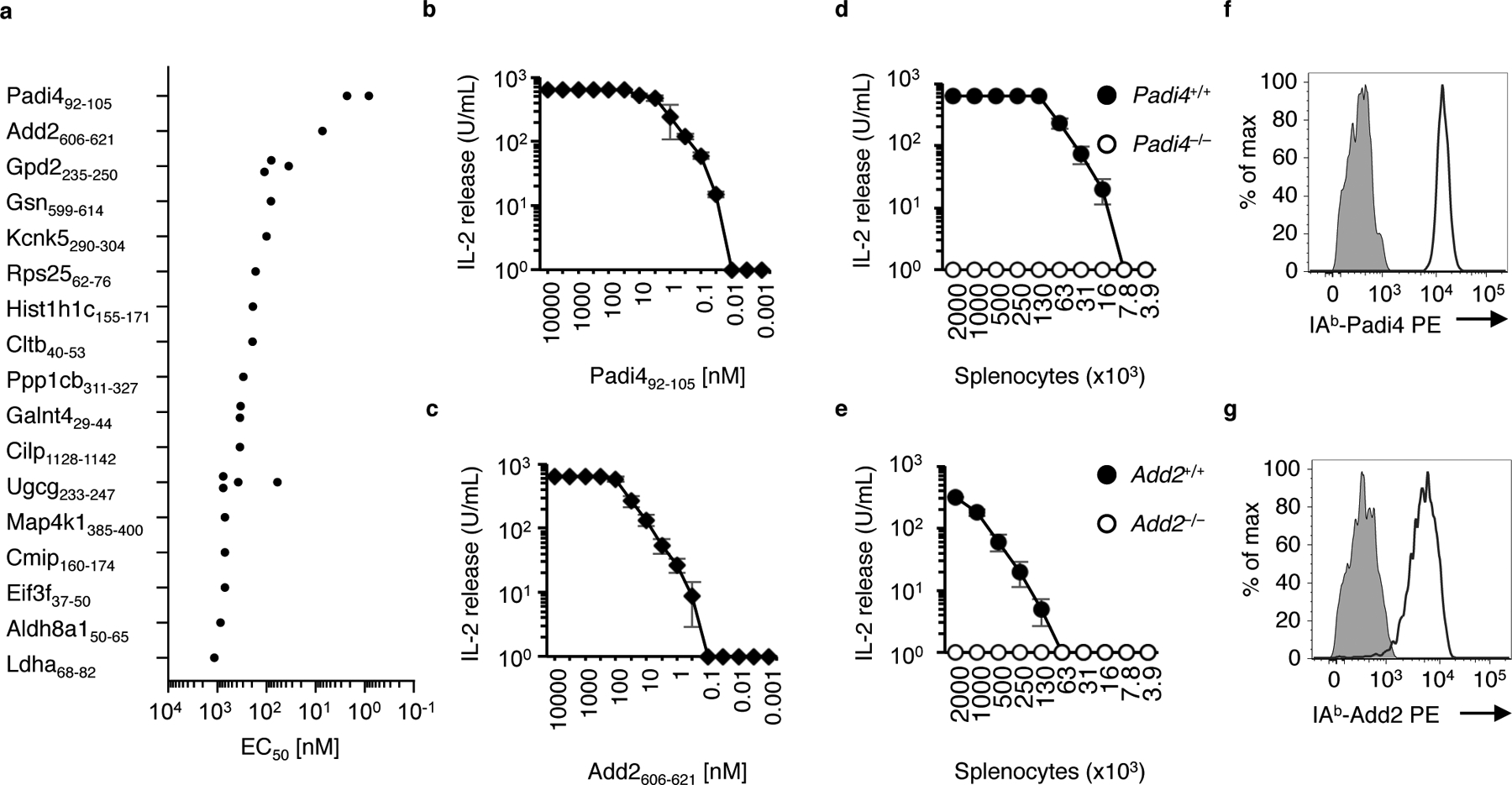
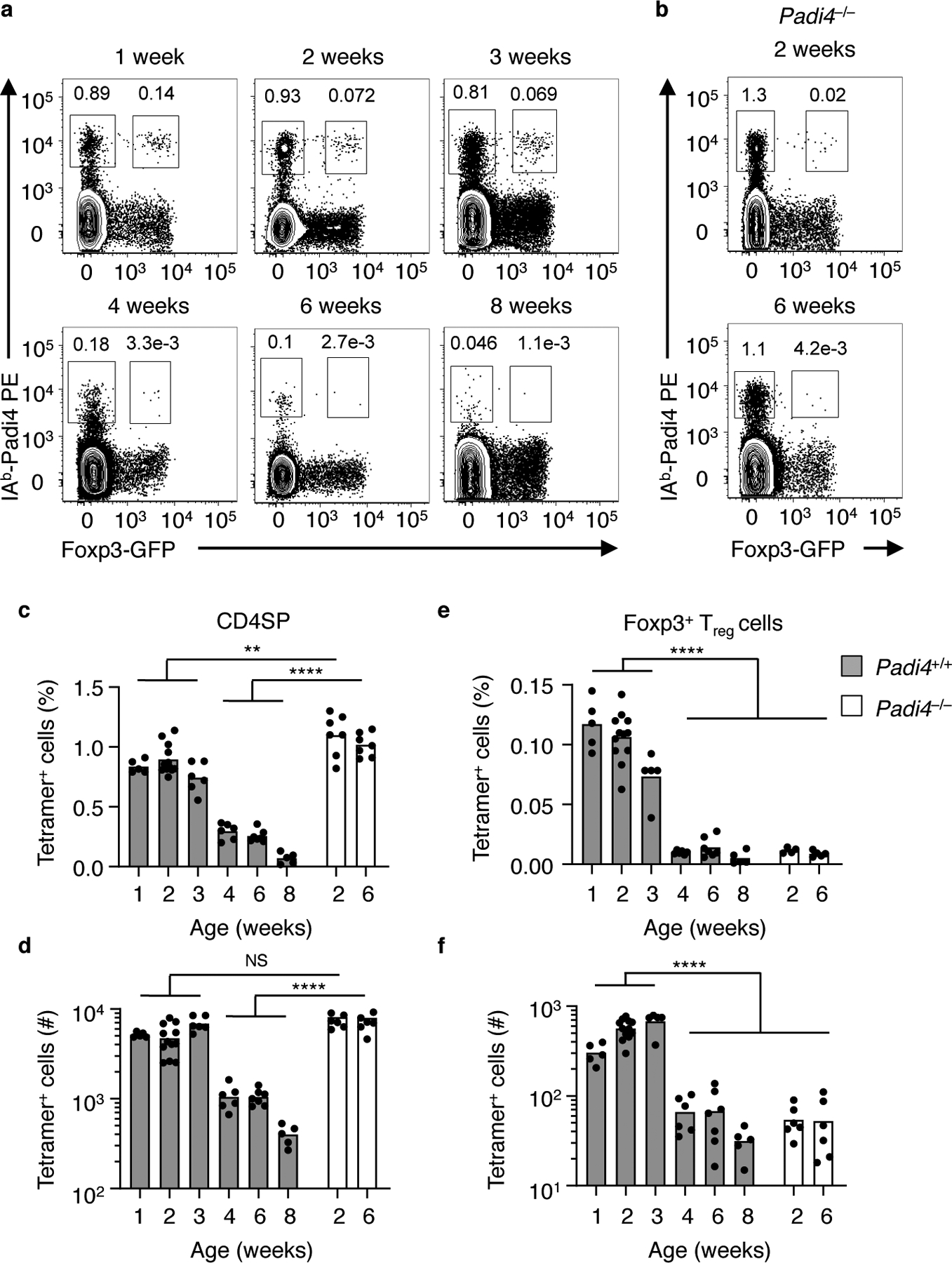
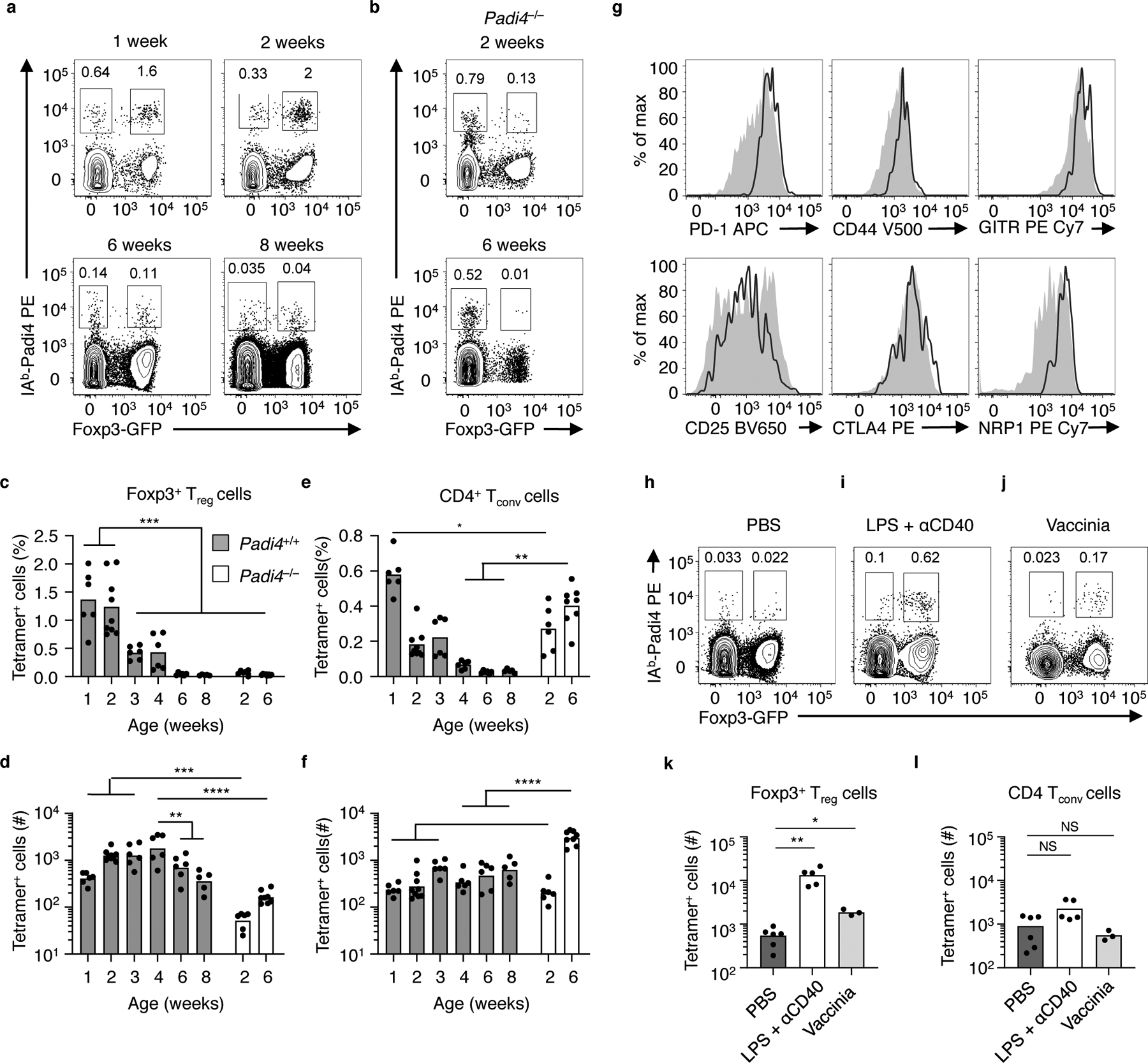
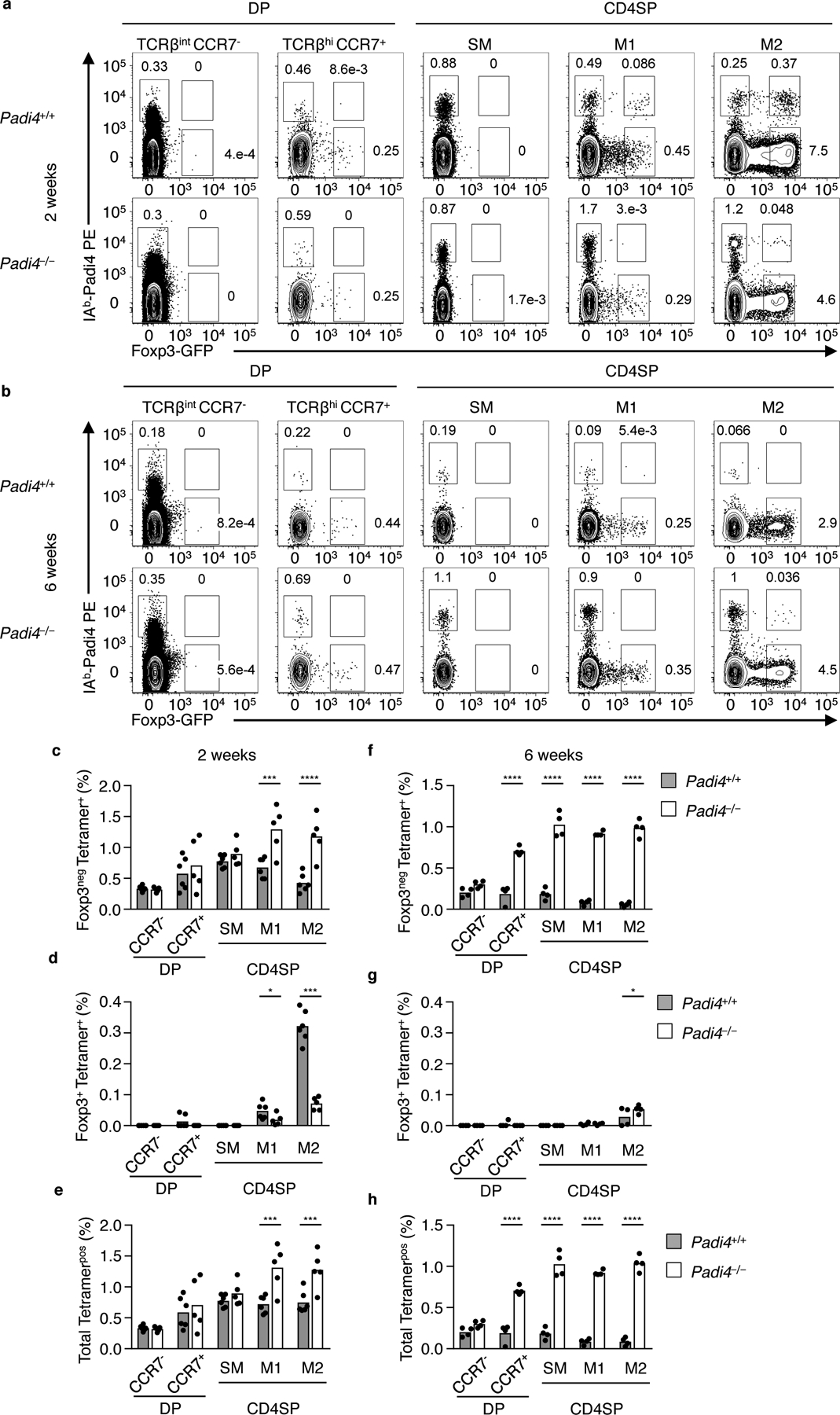
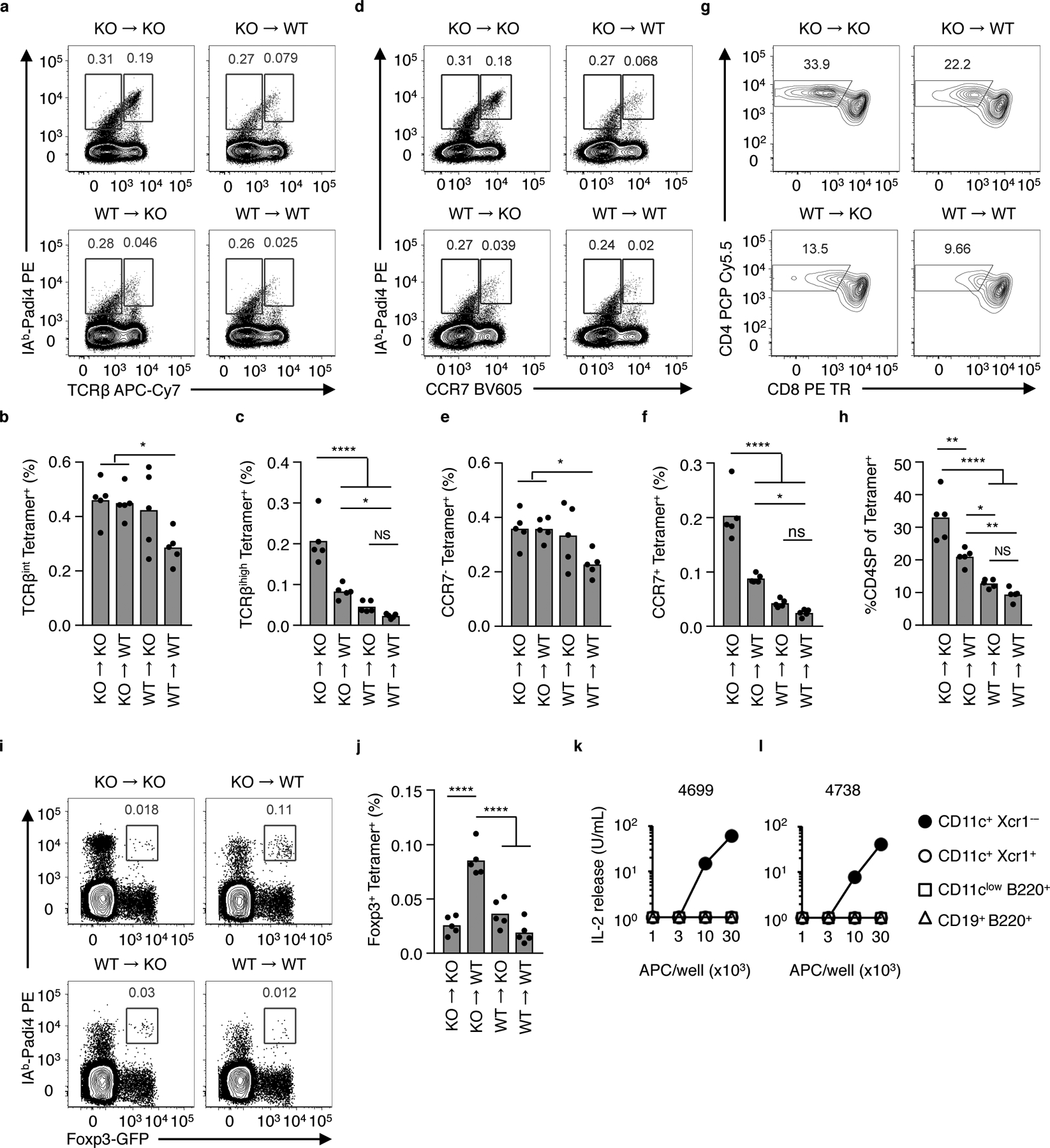
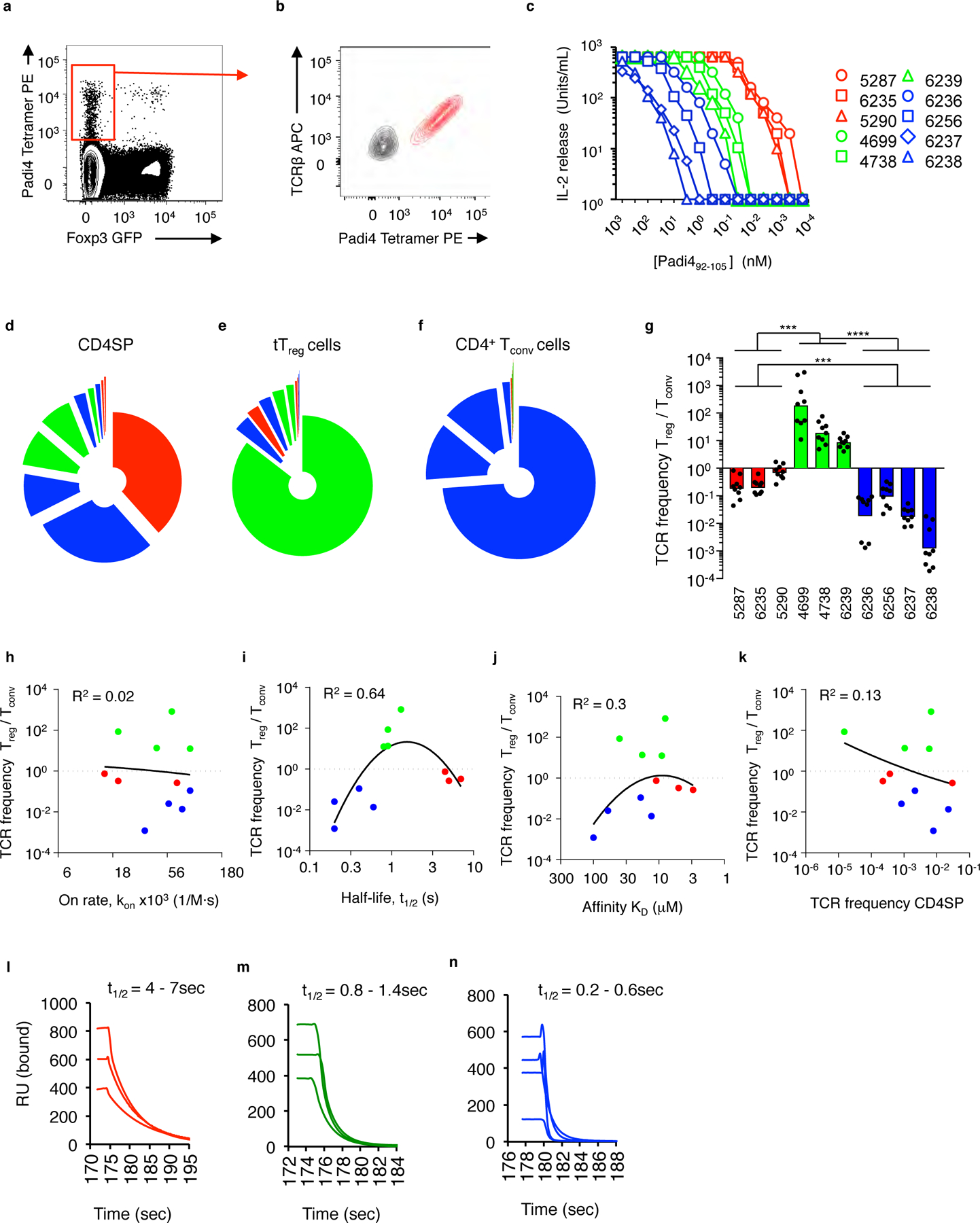
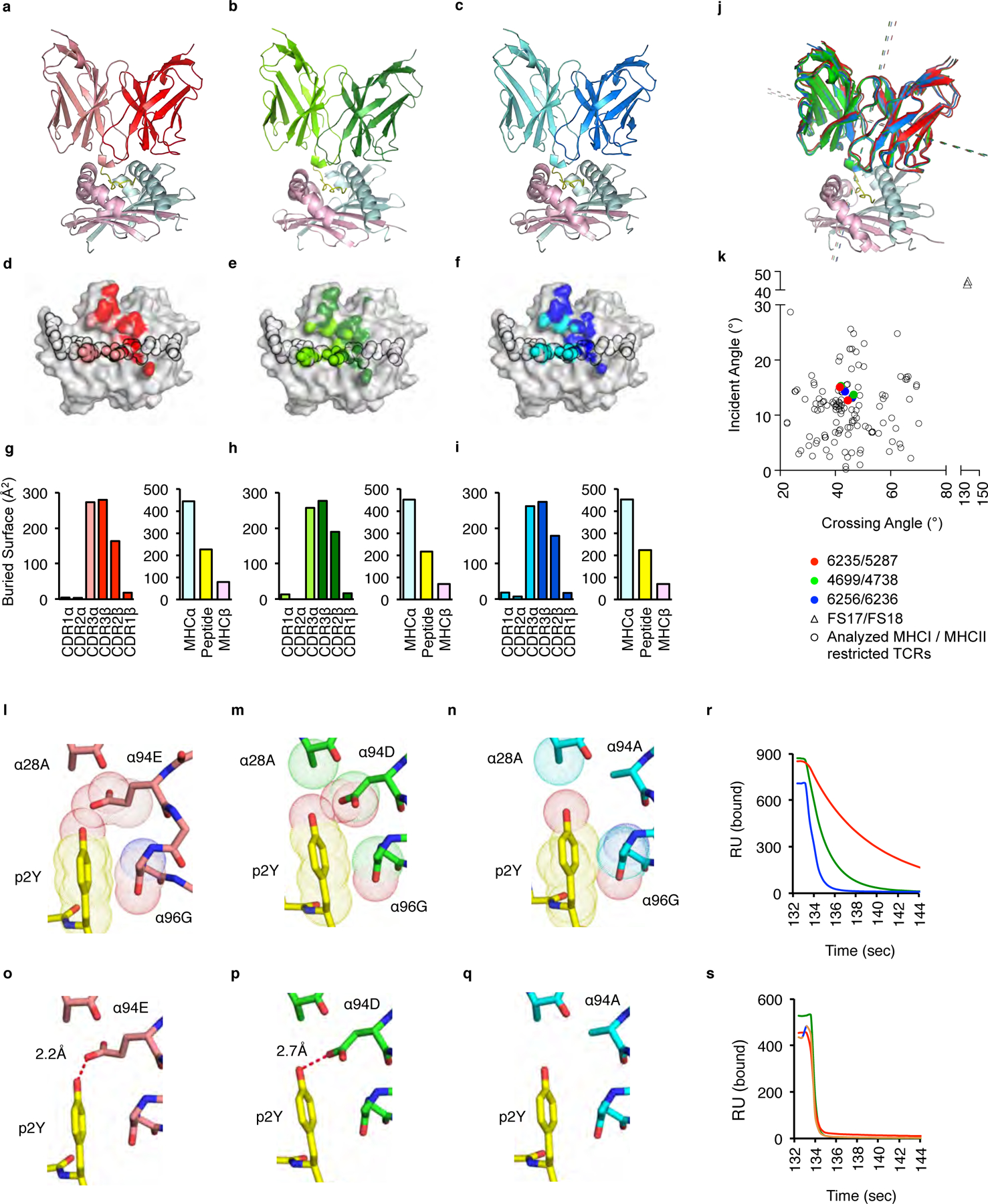
The neonatal thymus generates Foxp3+ regulatory T (tTreg) cells that are critical in controlling immune homeostasis and preventing multiorgan autoimmunity. The role of antigen specificity on neonatal tTreg cell selection is unresolved. Here we identify 17 self-peptides recognized by neonatal tTreg cells, and reveal ligand specificity patterns that include self-antigens presented in an age- and inflammation-dependent manner. Fate-mapping studies of neonatal peptidyl arginine deiminase type IV (Padi4)-specific thymocytes reveal disparate fate choices. Neonatal thymocytes expressing T cell receptors that engage IAb-Padi4 with moderate dwell times within a conventional docking orientation are exported as tTreg cells. In contrast, Padi4-specific T cell receptors with short dwell times are expressed on CD4+ T cells, while long dwell times induce negative selection. Temporally, Padi4-specific thymocytes are subject to a developmental stage-specific change in negative selection, which precludes tTreg cell development. Thus, a temporal switch in negative selection and ligand binding kinetics constrains the neonatal tTreg selection window.
Conflict of interest statement
Competing interests
The authors declare no competing interests
Figures
Similar articles
-
Development of thymic Foxp3(+) regulatory T cells: TGF-β matters.Eur J Immunol. 2015 Apr;45(4):958-65. doi: 10.1002/eji.201444999. Epub 2015 Mar 18. Eur J Immunol. 2015. PMID: 25684698 Free PMC article. Review.
-
Selection of Foxp3+ regulatory T cells specific for self antigen expressed and presented by Aire+ medullary thymic epithelial cells.Nat Immunol. 2007 Apr;8(4):351-8. doi: 10.1038/ni1444. Epub 2007 Feb 25. Nat Immunol. 2007. PMID: 17322887
-
Capacity of tTreg generation is not impaired in the atrophied thymus.PLoS Biol. 2017 Nov 8;15(11):e2003352. doi: 10.1371/journal.pbio.2003352. eCollection 2017 Nov. PLoS Biol. 2017. PMID: 29117183 Free PMC article.
-
Defective selection of thymic regulatory T cells accompanies autoimmunity and pulmonary infiltrates in Tcra-deficient mice double transgenic for human La/Sjögren's syndrome-B and human La-specific TCR.J Immunol. 2015 Feb 15;194(4):1514-22. doi: 10.4049/jimmunol.1400319. Epub 2015 Jan 12. J Immunol. 2015. PMID: 25582858 Free PMC article.
-
Thymic commitment of regulatory T cells is a pathway of TCR-dependent selection that isolates repertoires undergoing positive or negative selection.Curr Top Microbiol Immunol. 2005;293:43-71. doi: 10.1007/3-540-27702-1_3. Curr Top Microbiol Immunol. 2005. PMID: 15981475 Review.
Cited by
-
Adjusting to self in the thymus: CD4 versus CD8 lineage commitment and regulatory T cell development.J Exp Med. 2024 Oct 7;221(10):e20230896. doi: 10.1084/jem.20230896. Epub 2024 Jul 9. J Exp Med. 2024. PMID: 38980291 Free PMC article. Review.
-
MHC Class II Presentation in Autoimmunity.Cells. 2023 Jan 14;12(2):314. doi: 10.3390/cells12020314. Cells. 2023. PMID: 36672249 Free PMC article. Review.
-
Relationship of 2D Affinity to T Cell Functional Outcomes.Int J Mol Sci. 2020 Oct 27;21(21):7969. doi: 10.3390/ijms21217969. Int J Mol Sci. 2020. PMID: 33120989 Free PMC article. Review.
-
Your Regulatory T Cells Are What You Eat: How Diet and Gut Microbiota Affect Regulatory T Cell Development.Front Nutr. 2022 Apr 20;9:878382. doi: 10.3389/fnut.2022.878382. eCollection 2022. Front Nutr. 2022. PMID: 35529463 Free PMC article. Review.
-
Covalent TCR-peptide-MHC interactions induce T cell activation and redirect T cell fate in the thymus.Nat Commun. 2022 Aug 23;13(1):4951. doi: 10.1038/s41467-022-32692-4. Nat Commun. 2022. PMID: 35999236 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
