

The nucleosome position-encoding WW/SS sequence pattern is depleted in mammalian genes relative to other eukaryotes
- PMID: 31216031
- PMCID: PMC6735720
- DOI: 10.1093/nar/gkz544
The nucleosome position-encoding WW/SS sequence pattern is depleted in mammalian genes relative to other eukaryotes
Abstract
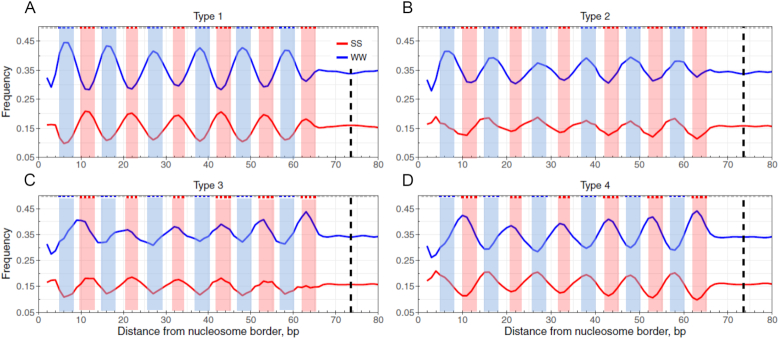
Nucleosomal DNA sequences generally follow a well-known pattern with ∼10-bp periodic WW (where W is A or T) dinucleotides that oscillate in phase with each other and out of phase with SS (where S is G or C) dinucleotides. However, nucleosomes with other DNA patterns have not been systematically analyzed. Here, we focus on an opposite pattern, namely anti-WW/SS pattern, in which WW dinucleotides preferentially occur at DNA sites that bend into major grooves and SS (where S is G or C) dinucleotides are often found at sites that bend into minor grooves. Nucleosomes with the anti-WW/SS pattern are widespread and exhibit a species- and context-specific distribution in eukaryotic genomes. Unlike non-mammals (yeast, nematode and fly), there is a positive correlation between the enrichment of anti-WW/SS nucleosomes and RNA Pol II transcriptional levels in mammals (mouse and human). Interestingly, such enrichment is not due to underlying DNA sequence. In addition, chromatin remodeling complexes have an impact on the abundance but not on the distribution of anti-WW/SS nucleosomes in yeast. Our data reveal distinct roles of cis- and trans-acting factors in the rotational positioning of nucleosomes between non-mammals and mammals. Implications of the anti-WW/SS sequence pattern for RNA Pol II transcription are discussed.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures








References
-
- Luger K., Mader A.W., Richmond R.K., Sargent D.F., Rochmond T.J.. Crystal structure of the nucleosome core particle at 2.8Å resolution. Nature. 1997; 389:251–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
