

Characterization of pathogenic monoclonal autoantibodies derived from muscle-specific kinase myasthenia gravis patients
- PMID: 31217355
- PMCID: PMC6629167
- DOI: 10.1172/jci.insight.127167
Characterization of pathogenic monoclonal autoantibodies derived from muscle-specific kinase myasthenia gravis patients
Abstract
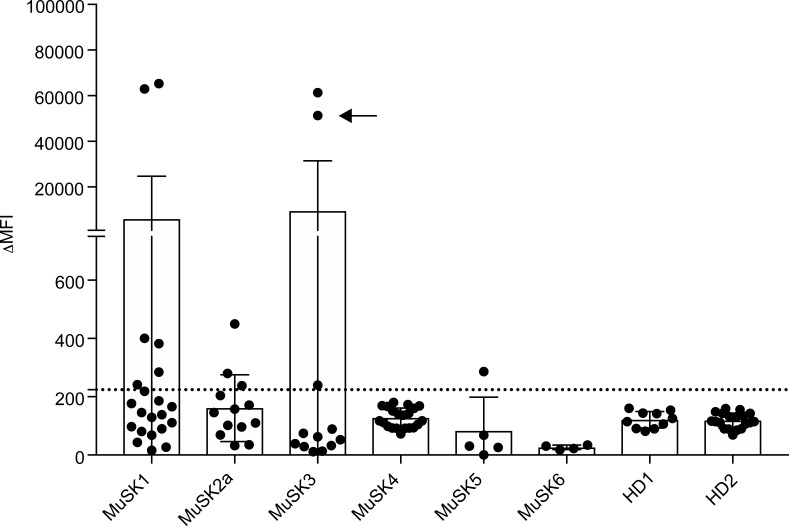
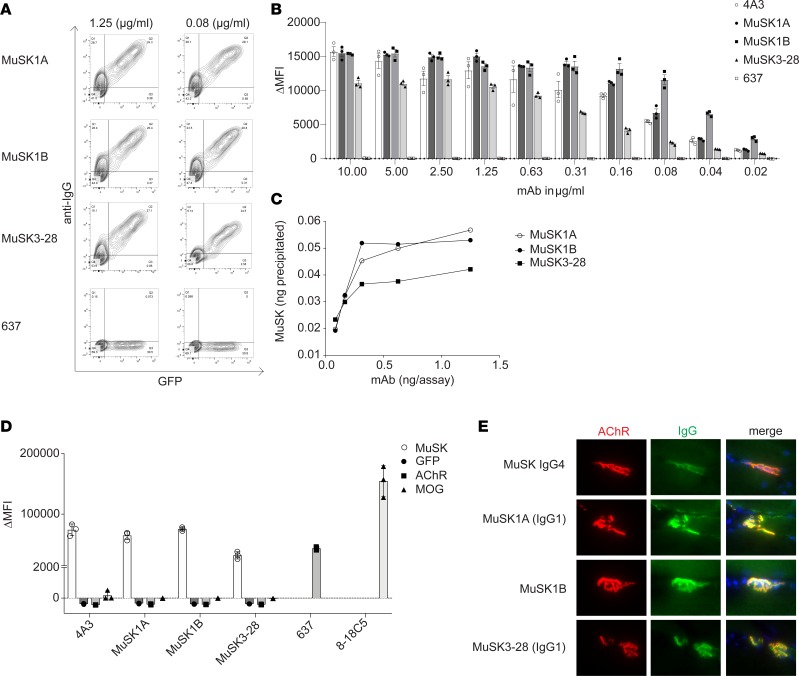
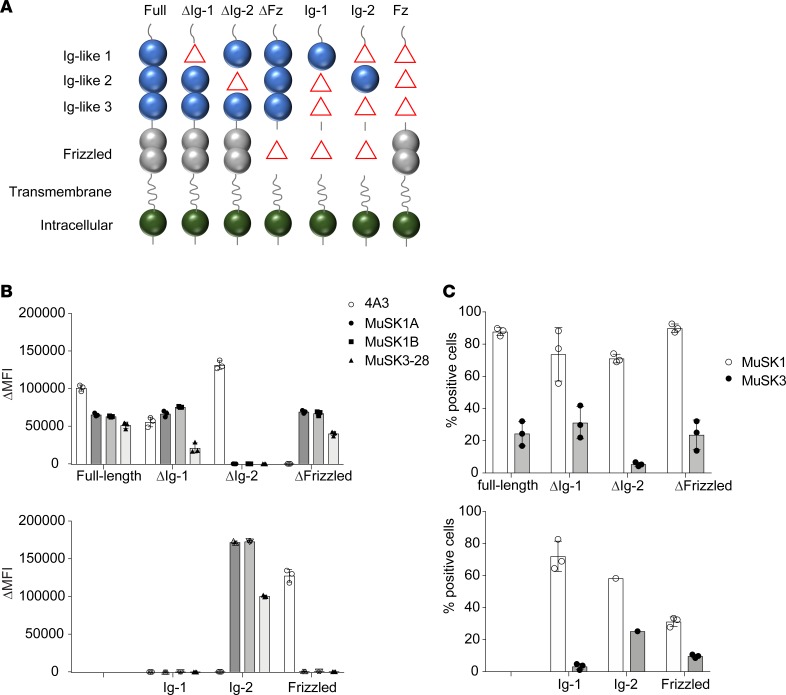
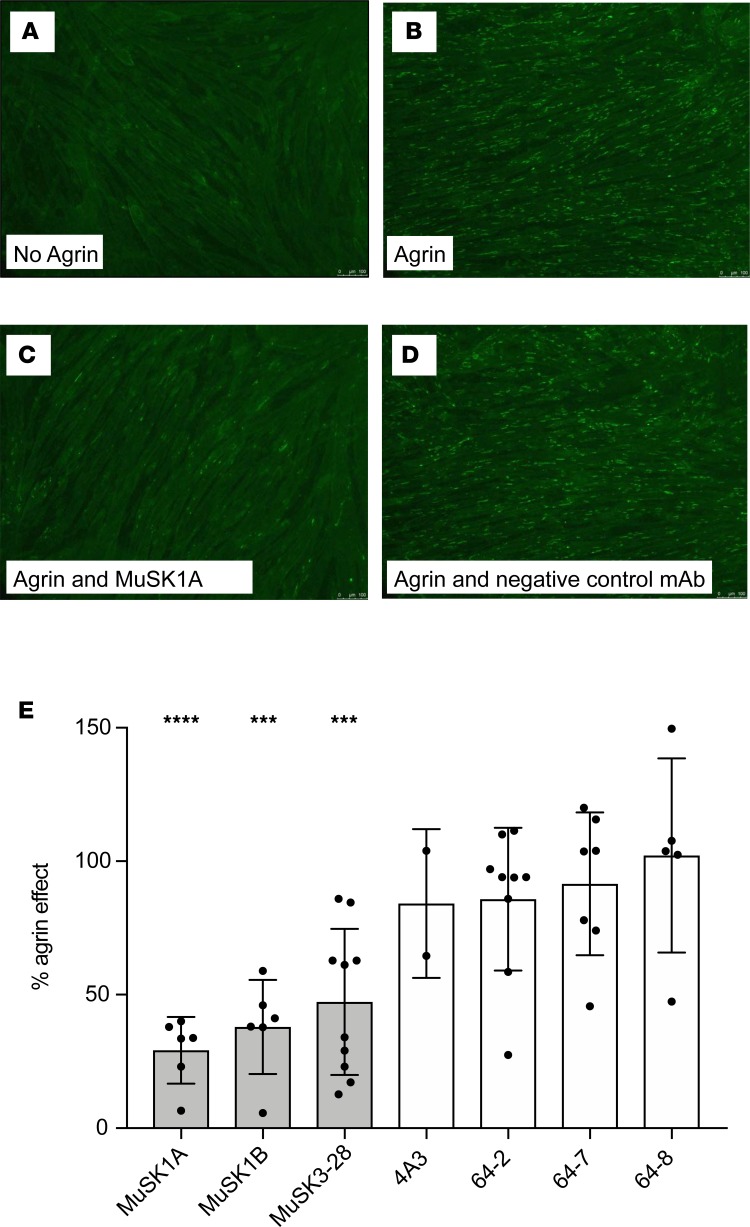
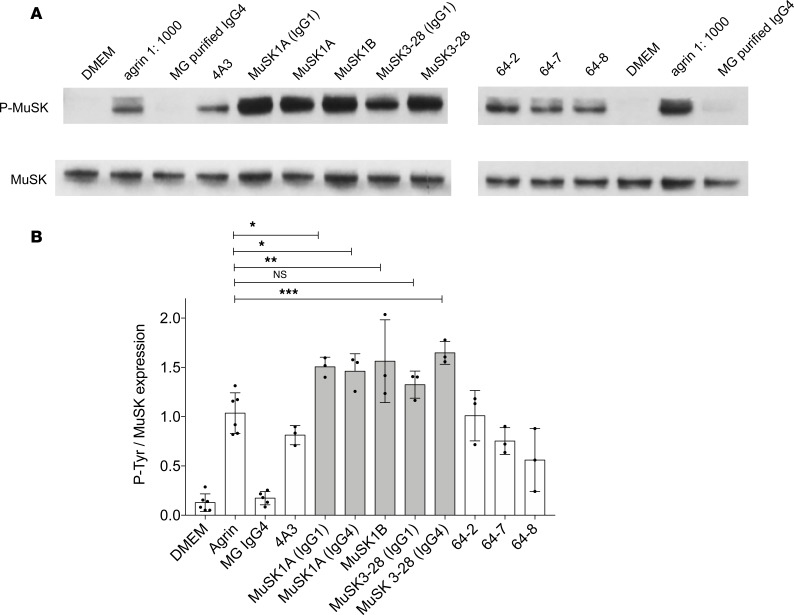
Myasthenia gravis (MG) is a chronic autoimmune disorder characterized by muscle weakness and caused by pathogenic autoantibodies that bind to membrane proteins at the neuromuscular junction. Most patients have autoantibodies against the acetylcholine receptor (AChR), but a subset of patients have autoantibodies against muscle-specific tyrosine kinase (MuSK) instead. MuSK is an essential component of the pathway responsible for synaptic differentiation, which is activated by nerve-released agrin. Through binding MuSK, serum-derived autoantibodies inhibit agrin-induced MuSK autophosphorylation, impair clustering of AChRs, and block neuromuscular transmission. We sought to establish individual MuSK autoantibody clones so that the autoimmune mechanisms could be better understood. We isolated MuSK autoantibody-expressing B cells from 6 MuSK MG patients using a fluorescently tagged MuSK antigen multimer, then generated a panel of human monoclonal autoantibodies (mAbs) from these cells. Here we focused on 3 highly specific mAbs that bound quantitatively to MuSK in solution, to MuSK-expressing HEK cells, and at mouse neuromuscular junctions, where they colocalized with AChRs. These 3 IgG isotype mAbs (2 IgG4 and 1 IgG3 subclass) recognized the Ig-like domain 2 of MuSK. The mAbs inhibited AChR clustering, but intriguingly, they enhanced rather than inhibited MuSK phosphorylation, which suggests an alternative mechanism for inhibiting AChR clustering.
Keywords: Autoimmune diseases; Autoimmunity; B cells; Immunology; Neuromuscular disease.
Conflict of interest statement
Figures
Similar articles
-
IgG4 autoantibodies against muscle-specific kinase undergo Fab-arm exchange in myasthenia gravis patients.J Autoimmun. 2017 Feb;77:104-115. doi: 10.1016/j.jaut.2016.11.005. Epub 2016 Dec 10. J Autoimmun. 2017. PMID: 27965060
-
MuSK myasthenia gravis monoclonal antibodies: Valency dictates pathogenicity.Neurol Neuroimmunol Neuroinflamm. 2019 Feb 21;6(3):e547. doi: 10.1212/NXI.0000000000000547. eCollection 2019 May. Neurol Neuroimmunol Neuroinflamm. 2019. PMID: 30882021 Free PMC article.
-
MuSK IgG4 autoantibodies cause myasthenia gravis by inhibiting binding between MuSK and Lrp4.Proc Natl Acad Sci U S A. 2013 Dec 17;110(51):20783-8. doi: 10.1073/pnas.1313944110. Epub 2013 Dec 2. Proc Natl Acad Sci U S A. 2013. PMID: 24297891 Free PMC article. Clinical Trial.
-
Antibody effector mechanisms in myasthenia gravis-pathogenesis at the neuromuscular junction.Autoimmunity. 2010 Aug;43(5-6):353-70. doi: 10.3109/08916930903555943. Autoimmunity. 2010. PMID: 20380584 Review.
-
Passive transfer models of myasthenia gravis with muscle-specific kinase antibodies.Ann N Y Acad Sci. 2018 Feb;1413(1):111-118. doi: 10.1111/nyas.13543. Epub 2018 Jan 21. Ann N Y Acad Sci. 2018. PMID: 29356029 Review.
Cited by
-
Complement Inhibitor Therapy for Myasthenia Gravis.Front Immunol. 2020 Jun 3;11:917. doi: 10.3389/fimmu.2020.00917. eCollection 2020. Front Immunol. 2020. PMID: 32582144 Free PMC article. Review.
-
Clinicoserological insights into patients with immune checkpoint inhibitor-induced myasthenia gravis.Ann Clin Transl Neurol. 2023 May;10(5):825-831. doi: 10.1002/acn3.51761. Epub 2023 Mar 16. Ann Clin Transl Neurol. 2023. PMID: 36924454 Free PMC article.
-
Efficacy and Safety of Low-Dose Rituximab in Anti-MuSK Myasthenia Gravis Patients: A Retrospective Study.Neuropsychiatr Dis Treat. 2022 May 3;18:953-964. doi: 10.2147/NDT.S358851. eCollection 2022. Neuropsychiatr Dis Treat. 2022. PMID: 35535211 Free PMC article.
-
Receptor clustering and pathogenic complement activation in myasthenia gravis depend on synergy between antibodies with multiple subunit specificities.Acta Neuropathol. 2022 Nov;144(5):1005-1025. doi: 10.1007/s00401-022-02493-6. Epub 2022 Sep 8. Acta Neuropathol. 2022. PMID: 36074148 Free PMC article.
-
Reemergence of pathogenic, autoantibody-producing B cell clones in myasthenia gravis following B cell depletion therapy.Acta Neuropathol Commun. 2022 Oct 28;10(1):154. doi: 10.1186/s40478-022-01454-0. Acta Neuropathol Commun. 2022. PMID: 36307868 Free PMC article.
References
-
- Lindstrom JM, Engel AG, Seybold ME, Lennon VA, Lambert EH. Pathological mechanisms in experimental autoimmune myasthenia gravis. II. Passive transfer of experimental autoimmune myasthenia gravis in rats with anti-acetylcholine receptor antibodies. J Exp Med. 1976;144(3):739–753. doi: 10.1084/jem.144.3.739. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
