

Cellular crosstalk in the development and regeneration of the respiratory system
- PMID: 31217577
- PMCID: PMC7254499
- DOI: 10.1038/s41580-019-0141-3
Cellular crosstalk in the development and regeneration of the respiratory system
Abstract
The respiratory system, including the peripheral lungs, large airways and trachea, is one of the most recently evolved adaptations to terrestrial life. To support the exchange of respiratory gases, the respiratory system is interconnected with the cardiovascular system, and this interconnective nature requires a complex interplay between a myriad of cell types. Until recently, this complexity has hampered our understanding of how the respiratory system develops and responds to postnatal injury to maintain homeostasis. The advent of new single-cell sequencing technologies, developments in cellular and tissue imaging and advances in cell lineage tracing have begun to fill this gap. The view that emerges from these studies is that cellular and functional heterogeneity of the respiratory system is even greater than expected and also highly adaptive. In this Review, we explore the cellular crosstalk that coordinates the development and regeneration of the respiratory system. We discuss both the classic cell and developmental biology studies and recent single-cell analysis to provide an integrated understanding of the cellular niches that control how the respiratory system develops, interacts with the external environment and responds to injury.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures

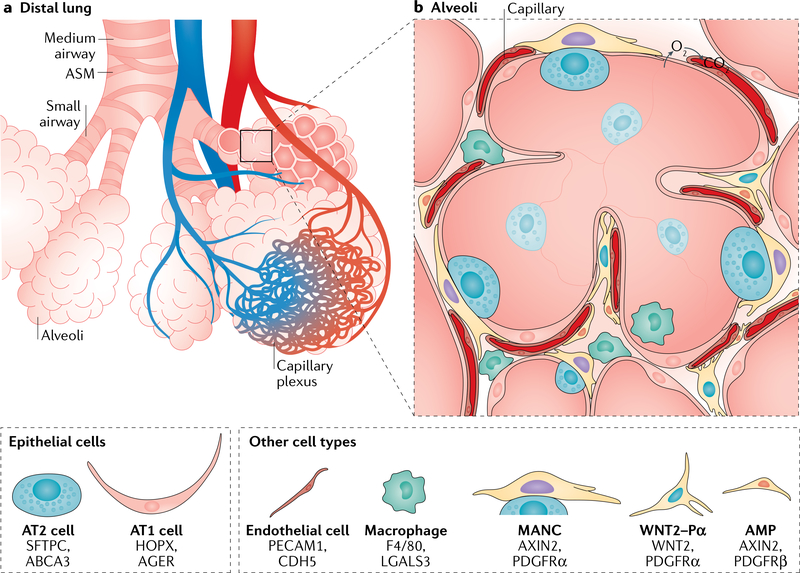

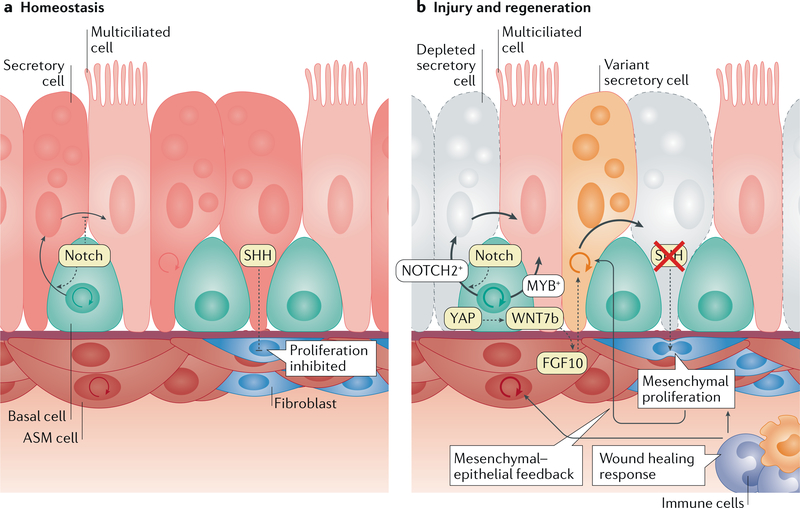
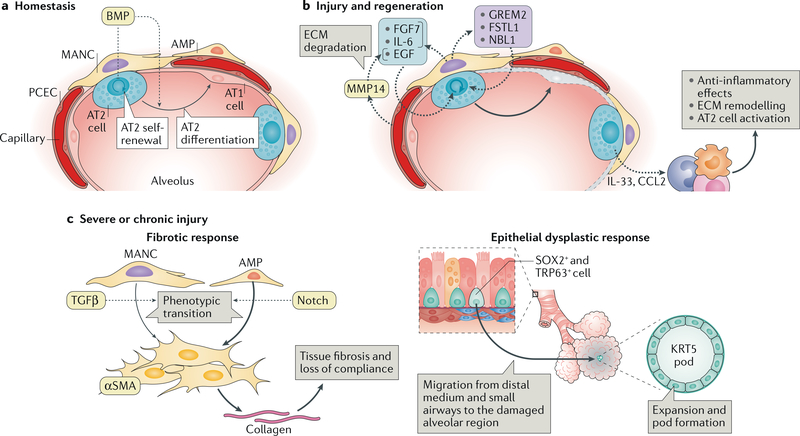
References
-
- Volckaert T et al. Parabronchial smooth muscle constitutes an airway epithelial stem cell niche in the mouse lung after injury. J. Clin. Invest. 121, 4409–4419 (2011). Lee et al. (2014) and Volckaert et al. (2011) demonstrate the requirement of anatomically distinct mesenchymal cell types, such as LGR6- positive airway smooth muscle cells in promoting epithelial recovery after injury. - PMC - PubMed
-
-
Barkauskas CE et al. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Invest. 123, 3025–3036 (2013).
This study uses genetic lineage tracing techniques to show that AT2 cells contain a progenitor cell function in the adult lung and can differentiate into AT1 cells.
-
-
- Caduff JH, Fischer LC & Burri PH Scanning electron microscope study of the developing microvasculature in the postnatal rat lung. Anat. Rec. 216, 154–164 (1986). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
