

FAT4-USP51 complex regulates the proliferation and invasion of endometrial cancer via Hippo pathway
- PMID: 31217854
- PMCID: PMC6556641
FAT4-USP51 complex regulates the proliferation and invasion of endometrial cancer via Hippo pathway
Abstract
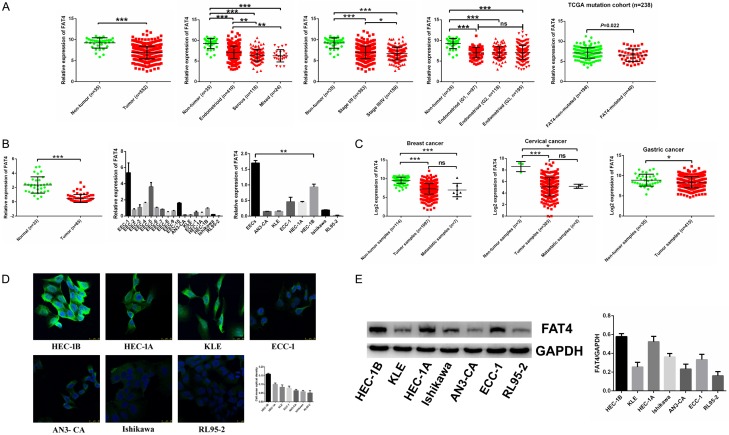
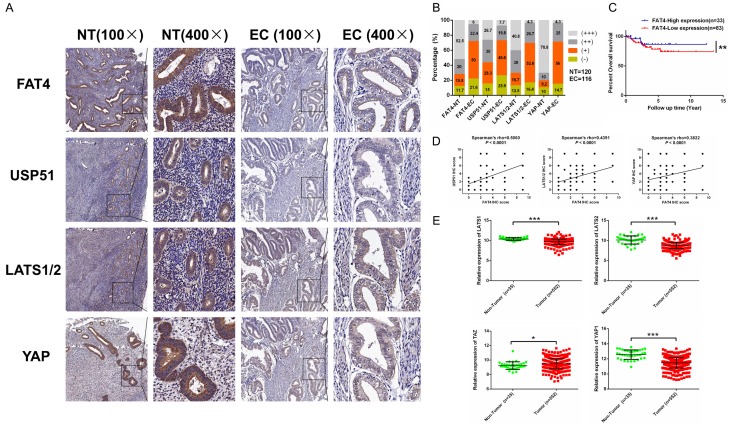
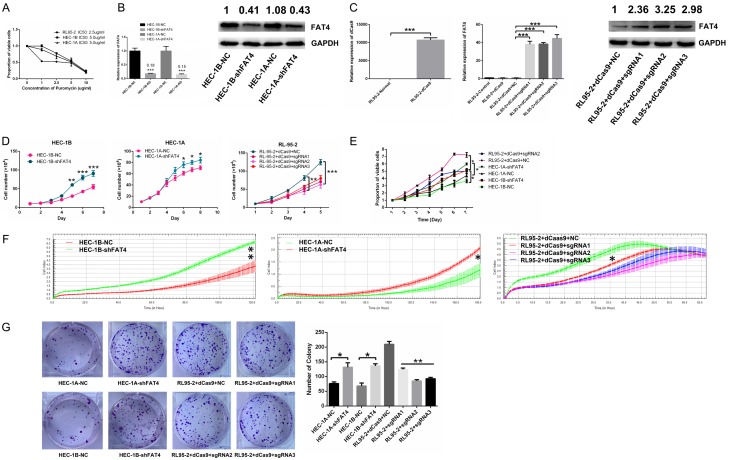
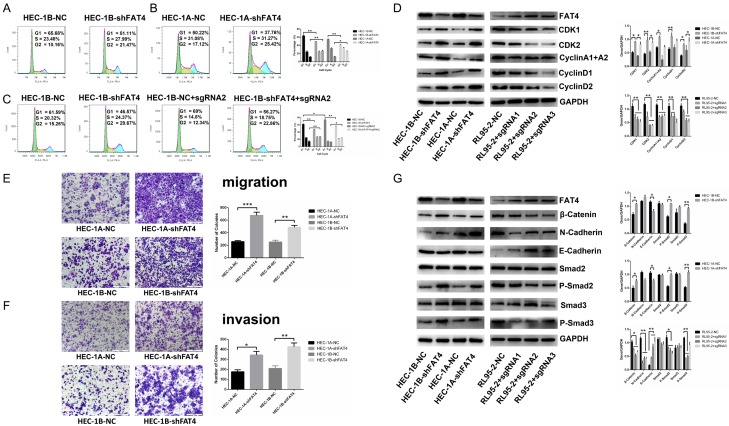
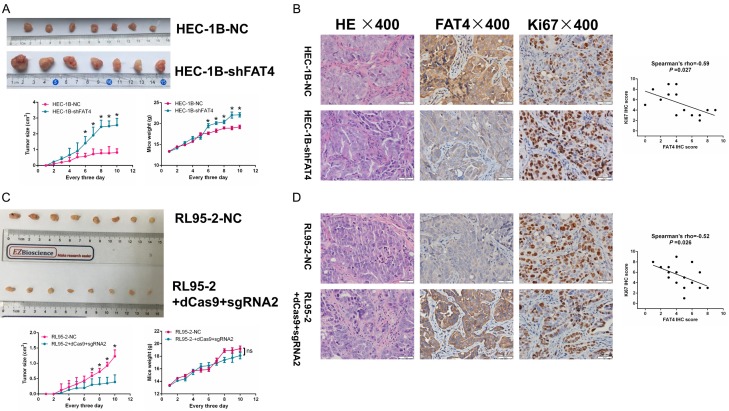
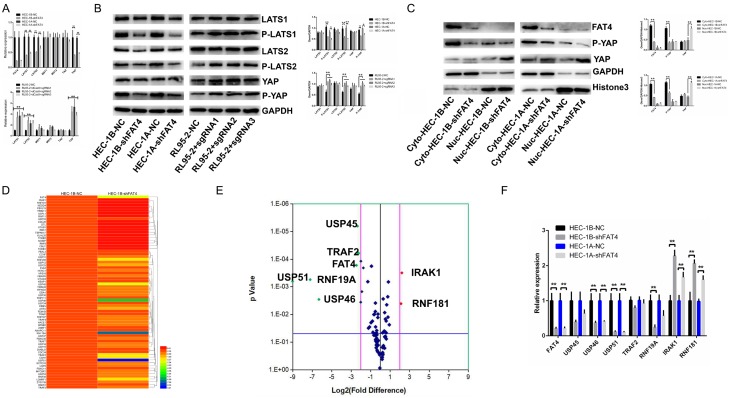
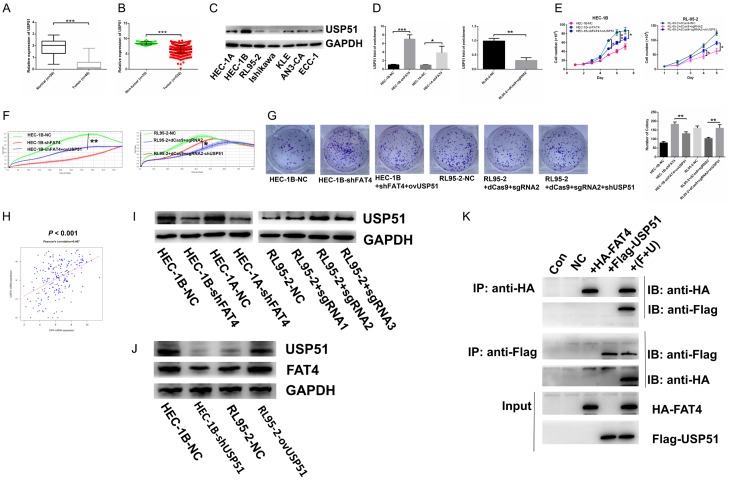
Recent studies have identified FAT tumour suppressor homologue 4 (FAT4), an essential component of adherents junctions, involved in several cancers. However, its role in endometrial cancer (EC) remains unclear. In this study, we first analyzed the association between FAT4 expression and tumour stage, tumour type, and patient prognosis in 552 tumour samples and 35 non-tumour samples from The Cancer Genome Atlas (TCGA) database. The association of decreased FAT4 expression with advanced signature (lymph node metastasis, lymphovascular invasion and muscular infiltration) in EC patients was also confirmed by our own dataset. Stable FAT4 Knockdown promoted EC cell lines proliferation and invasion. FAT4 overexpression inhibited the parental cell phenotype. FAT4 silencing resulted in decreased phosphorylation of the LATS1/2 and YAP while increased YAP nuclear translocation which was associated with the promotion of proliferation and invasion. PCR array analysis of the negative control and shFAT4 HEC-1B cell lines revealed that the deubiquitinating enzyme USP51 was a FAT4 interacting target gene. Ablating USP51 by shRNA decreased cellular FAT4 protein level while overexpression of USP51 increased FAT4 protein level. Coimmunoprecipitation confirmed the direct binding of FAT4 and USP51 which was essential for FAT4's function in EC. The growth inhibitory effect of FAT4 was also attenuated by USP51 down-regulation. In conclusion, suppression of FAT4 by inactivation of deubiquitinating enzyme USP51 promoted proliferation and invasion of EC cells via inhibiting Hippo pathway.
Keywords: FAT4; Hippo pathway; USP51; endometrial cancer.
Conflict of interest statement
None.
Figures
Similar articles
-
Fat4 suppression induces Yap translocation accounting for the promoted proliferation and migration of gastric cancer cells.Cancer Biol Ther. 2016;17(1):36-47. doi: 10.1080/15384047.2015.1108488. Cancer Biol Ther. 2016. PMID: 26575609 Free PMC article.
-
The novel FAT4 activator jujuboside A suppresses NSCLC tumorigenesis by activating HIPPO signaling and inhibiting YAP nuclear translocation.Pharmacol Res. 2021 Aug;170:105723. doi: 10.1016/j.phrs.2021.105723. Epub 2021 Jun 9. Pharmacol Res. 2021. PMID: 34116210
-
FAT4 functions as a tumour suppressor in gastric cancer by modulating Wnt/β-catenin signalling.Br J Cancer. 2015 Dec 22;113(12):1720-9. doi: 10.1038/bjc.2015.367. Epub 2015 Dec 3. Br J Cancer. 2015. PMID: 26633557 Free PMC article.
-
WWC3 regulates the Wnt and Hippo pathways via Dishevelled proteins and large tumour suppressor 1, to suppress lung cancer invasion and metastasis.J Pathol. 2017 Aug;242(4):435-447. doi: 10.1002/path.4919. Epub 2017 Jun 29. J Pathol. 2017. PMID: 28543074
-
Function and cancer genomics of FAT family genes (review).Int J Oncol. 2012 Dec;41(6):1913-8. doi: 10.3892/ijo.2012.1669. Epub 2012 Oct 17. Int J Oncol. 2012. PMID: 23076869 Free PMC article. Review.
Cited by
-
FAT4 overexpression promotes antitumor immunity by regulating the β-catenin/STT3/PD-L1 axis in cervical cancer.J Exp Clin Cancer Res. 2023 Sep 1;42(1):222. doi: 10.1186/s13046-023-02758-2. J Exp Clin Cancer Res. 2023. PMID: 37658376 Free PMC article.
-
MiR-106b-5p regulates the migration and invasion of colorectal cancer cells by targeting FAT4.Biosci Rep. 2020 Nov 27;40(11):BSR20200098. doi: 10.1042/BSR20200098. Biosci Rep. 2020. PMID: 33063118 Free PMC article.
-
Identification of FAT4 as a positive prognostic biomarker in DLBCL by comprehensive genomic analysis.Clin Exp Med. 2023 Oct;23(6):2675-2685. doi: 10.1007/s10238-023-01018-z. Epub 2023 Feb 22. Clin Exp Med. 2023. PMID: 36811800 Free PMC article.
-
Increased expression of FAT4 suppress metastasis of lung adenocarcinoma through regulating MAPK pathway and associated with immune cells infiltration.Cancer Med. 2023 Jan;12(2):1616-1629. doi: 10.1002/cam4.4977. Epub 2022 Jun 30. Cancer Med. 2023. PMID: 35770846 Free PMC article.
-
Deubiquitylating Enzymes in Cancer and Immunity.Adv Sci (Weinh). 2023 Dec;10(36):e2303807. doi: 10.1002/advs.202303807. Epub 2023 Oct 27. Adv Sci (Weinh). 2023. PMID: 37888853 Free PMC article. Review.
References
-
- Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66:115–32. - PubMed
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68:7–30. - PubMed
-
- Bokhman JV. Two pathogenetic types of endometrial carcinoma. Gynecol Oncol. 1983;15:10–7. - PubMed
-
- Setiawan VW, Yang HP, Pike MC, McCann SE, Yu H, Xiang YB, Wolk A, Wentzensen N, Weiss NS, Webb PM, van den Brandt PA, van de Vijver K, Thompson PJ, Strom BL, Spurdle AB, Soslow RA, Shu XO, Schairer C, Sacerdote C, Rohan TE, Robien K, Risch HA, Ricceri F, Rebbeck TR, Rastogi R, Prescott J, Polidoro S, Park Y, Olson SH, Moysich KB, Miller AB, McCullough ML, Matsuno RK, Magliocco AM, Lurie G, Lu L, Lissowska J, Liang X, Lacey JJ, Kolonel LN, Henderson BE, Hankinson SE, Hakansson N, Goodman MT, Gaudet MM, Garcia-Closas M, Friedenreich CM, Freudenheim JL, Doherty J, De Vivo I, Courneya KS, Cook LS, Chen C, Cerhan JR, Cai H, Brinton LA, Bernstein L, Anderson KE, Anton-Culver H, Schouten LJ, Horn-Ross PL. Type I and II endometrial cancers: have they different risk factors? J Clin Oncol. 2013;31:2607–2618. - PMC - PubMed
-
- Hoeng JC, Ivanov NV, Hodor P, Xia M, Wei N, Blevins R, Gerhold D, Borodovsky M, Liu Y. Identification of new human cadherin genes using a combination of protein motif search and gene finding methods. J Mol Biol. 2004;337:307–317. - PubMed
LinkOut - more resources
Full Text Sources