

Metabolic Responses to Reductive Stress
- PMID: 31218894
- PMCID: PMC7247050
- DOI: 10.1089/ars.2019.7803
Metabolic Responses to Reductive Stress
Abstract
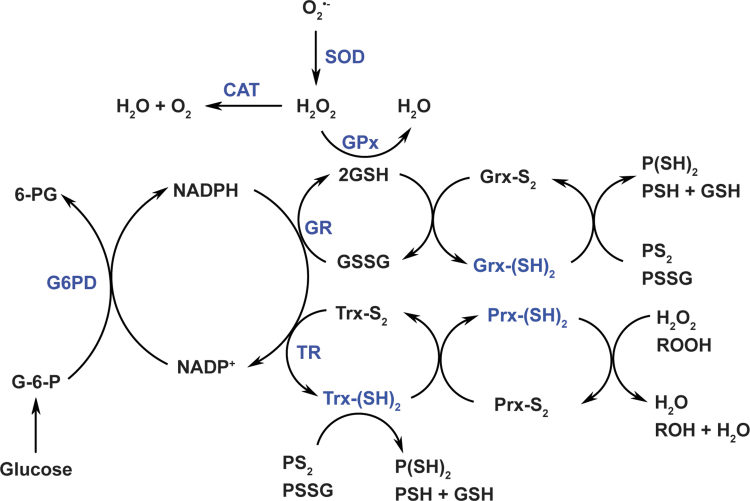
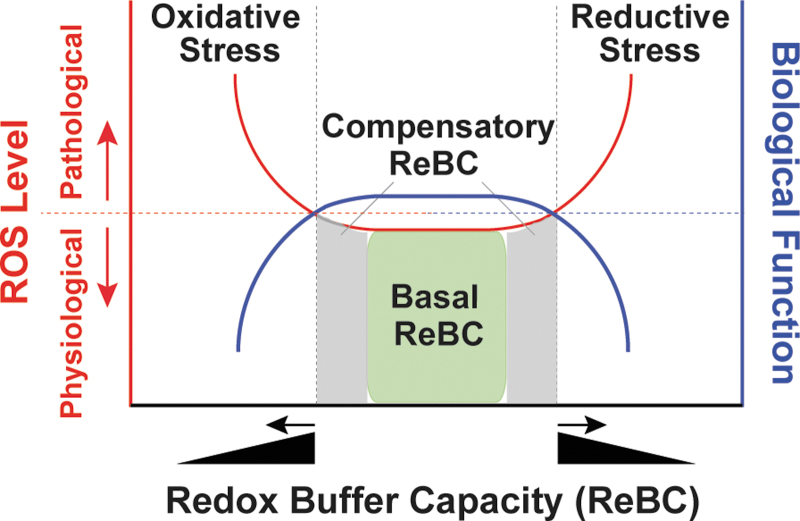
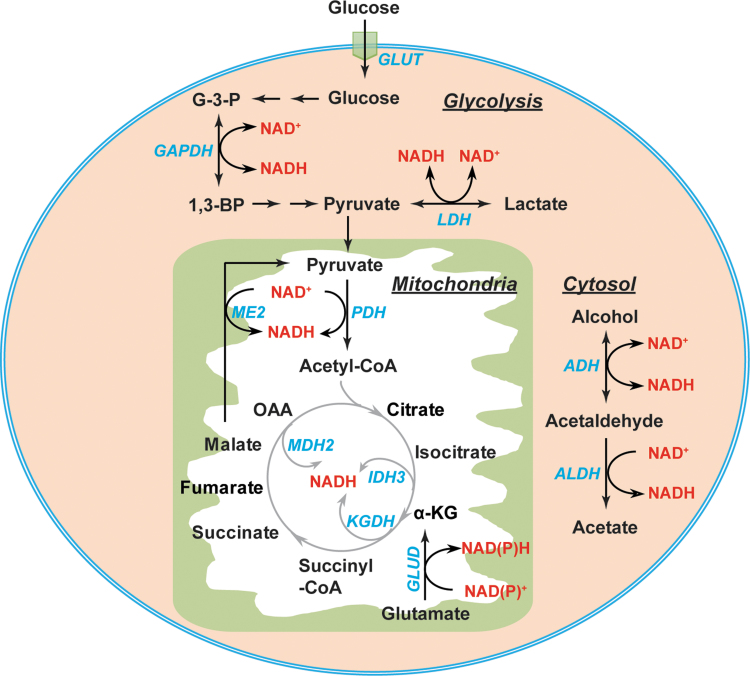
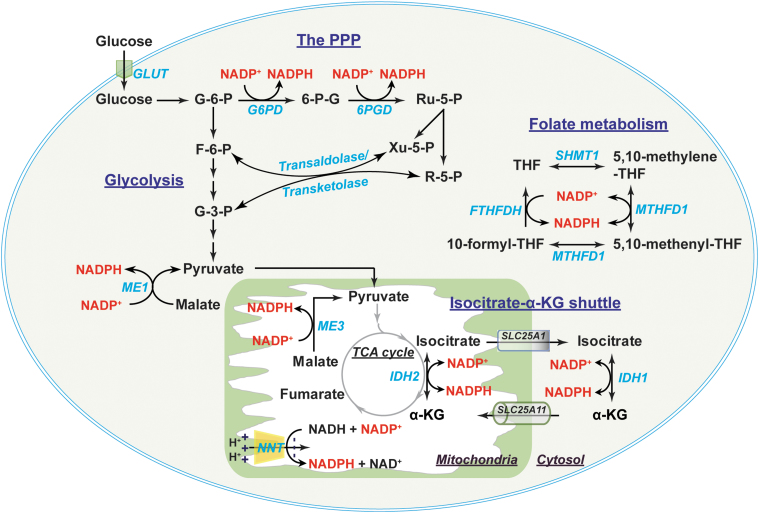
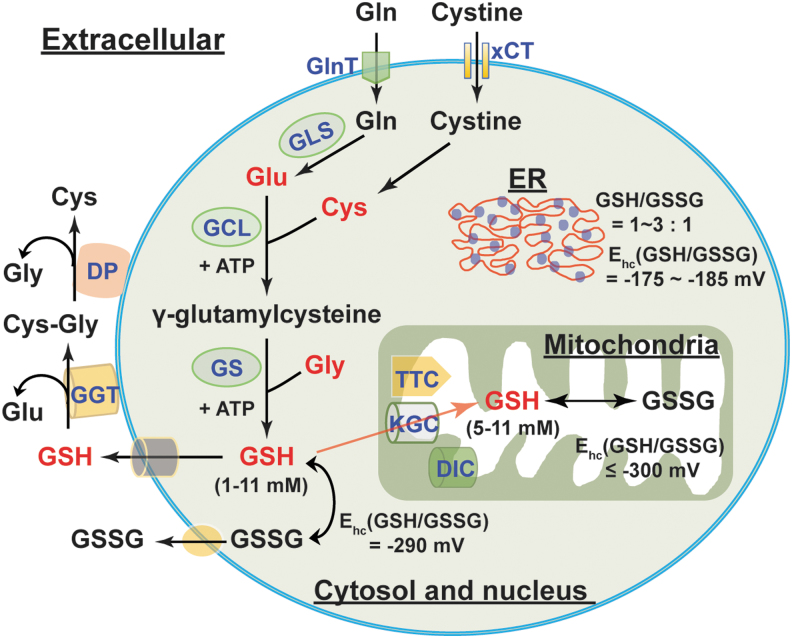
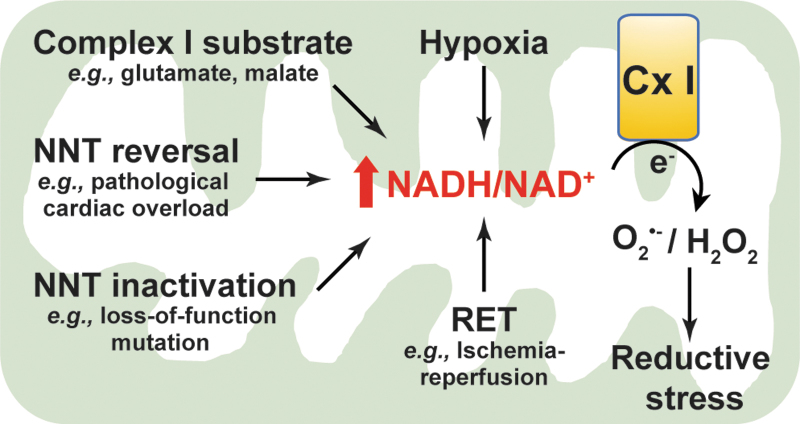
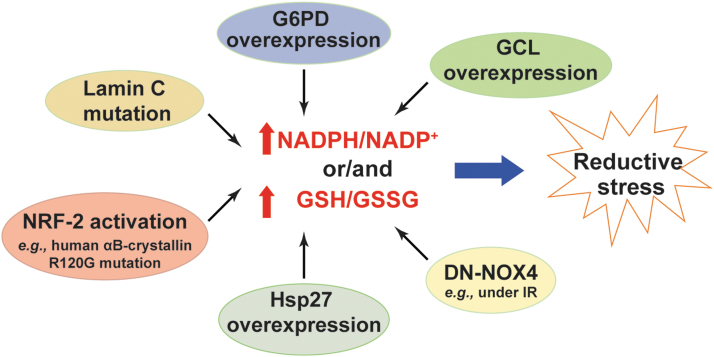
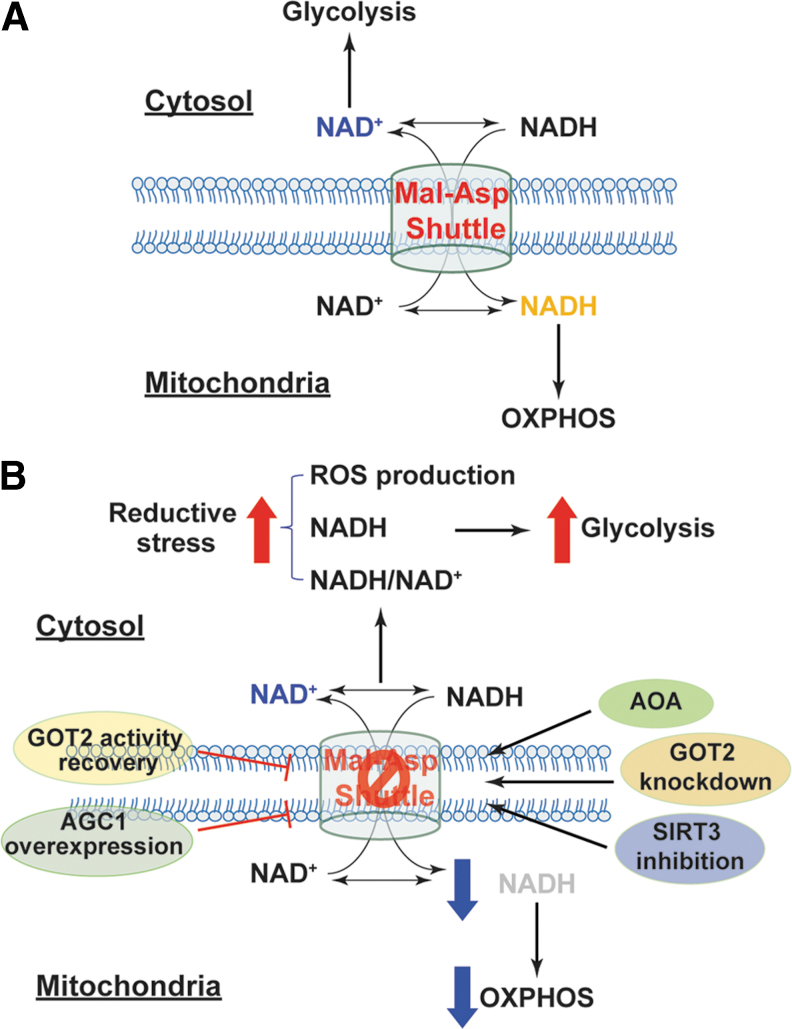
Significance: Reducing equivalents (NAD(P)H and glutathione [GSH]) are essential for maintaining cellular redox homeostasis and for modulating cellular metabolism. Reductive stress induced by excessive levels of reduced NAD+ (NADH), reduced NADP+ (NADPH), and GSH is as harmful as oxidative stress and is implicated in many pathological processes. Recent Advances: Reductive stress broadens our view of the importance of cellular redox homeostasis and the influences of an imbalanced redox niche on biological functions, including cell metabolism. Critical Issues: The distribution of cellular NAD(H), NADP(H), and GSH/GSH disulfide is highly compartmentalized. Understanding how cells coordinate different pools of redox couples under unstressed and stressed conditions is critical for a comprehensive view of redox homeostasis and stress. It is also critical to explore the underlying mechanisms of reductive stress and its biological consequences, including effects on energy metabolism. Future Directions: Future studies are needed to investigate how reductive stress affects cell metabolism and how cells adapt their metabolism to reductive stress. Whether or not NADH shuttles and mitochondrial nicotinamide nucleotide transhydrogenase enzyme can regulate hypoxia-induced reductive stress is also a worthy pursuit. Developing strategies (e.g., antireductant approaches) to counteract reductive stress and its related adverse biological consequences also requires extensive future efforts.
Keywords: GSH/GSSG; NAD(H); NADP(H); cellular metabolism; redox; reductive stress.
Figures
References
-
- Alderson NL, Wang Y, Blatnik M, Frizzell N, Walla MD, Lyons TJ, Alt N, Carson JA, Nagai R, Thorpe SR, and Baynes JW. S-(2-Succinyl)cysteine: a novel chemical modification of tissue proteins by a Krebs cycle intermediate. Arch Biochem Biophys 450: 1–8, 2006 - PubMed
-
- Barron JT, Gu L, and Parrillo JE. Malate-aspartate shuttle, cytoplasmic NADH redox potential, and energetics in vascular smooth muscle. J Mol Cell Cardiol 30: 1571–1579, 1998 - PubMed
-
- Barron JT, Gu L, and Parrillo JE. NADH/NAD redox state of cytoplasmic glycolytic compartments in vascular smooth muscle. Am J Physiol Heart Circ Physiol 279: H2872–H2878, 2000 - PubMed
-
- Bedard K and Krause KH. The NOX family of ROS-generating NADPH oxidases: physiology and pathophysiology. Physiol Rev 87: 245–313, 2007 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
