

Review
doi: 10.1093/femsre/fuz010.
Revisiting gametocyte biology in malaria parasites
Affiliations
- PMID: 31220244
- PMCID: PMC6606849
- DOI: 10.1093/femsre/fuz010
Item in Clipboard
Review
Revisiting gametocyte biology in malaria parasites
FEMS Microbiol Rev.
.
Erratum in
-
Corrigendum to: Revisiting gametocyte biology in malaria parasites.FEMS Microbiol Rev. 2019 Nov 1;43(6):684. doi: 10.1093/femsre/fuz020. FEMS Microbiol Rev. 2019. PMID: 31386107 Free PMC article. No abstract available.
Abstract
Gametocytes are the only form of the malaria parasite that is transmissible to the mosquito vector. They are present at low levels in blood circulation and significant knowledge gaps exist in their biology. Recent reductions in the global malaria burden have brought the possibility of elimination and eradication, with renewed focus on malaria transmission biology as a basis for interventions. This review discusses recent insights into gametocyte biology in the major human malaria parasite, Plasmodium falciparum and related species.
Keywords: Plasmodium falciparum; gametocyte; malaria; transmission.
© FEMS 2019.
Figures
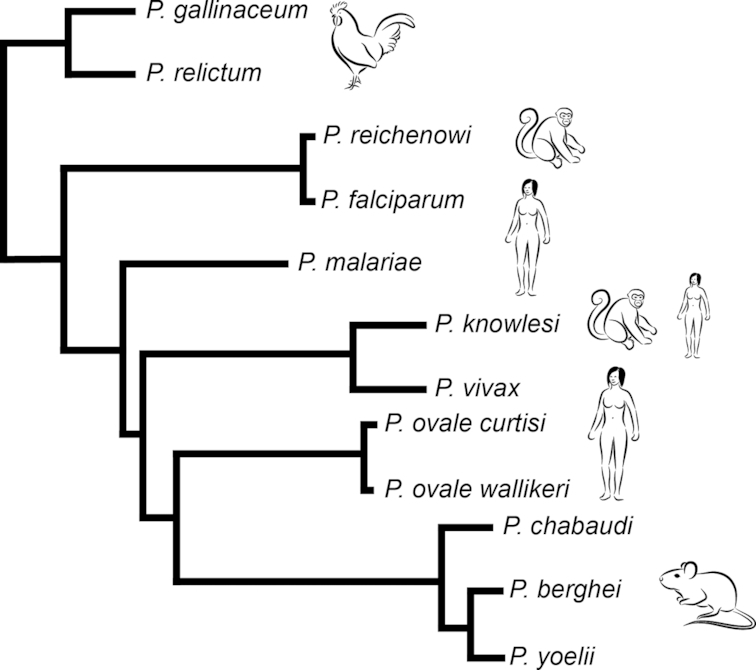
Phylogenetic representation of Plasmodium lineages. Schematic of phylogenetic tree based on most recent genome information across Plasmodium lineage. The tree is adapted from the recent publication by Boehme et al. (2018). The data show the major Plasmodium lineages: bird (P. gallinaceum, P. relictum) parasites are at the base of the tree whilst Laverania (including P. falciparum and P. reichenowi) are a sister group to other Plasmodium within the clade of mammalian parasites. The latter include rodent malaria parasites (P. berghei, P. chabaudi, P. yoelii) as well as primate (P. knowlesi) and human parasites (P. ovale curtisi and P. ovale wallikeri, P. vivax andP. malariae).
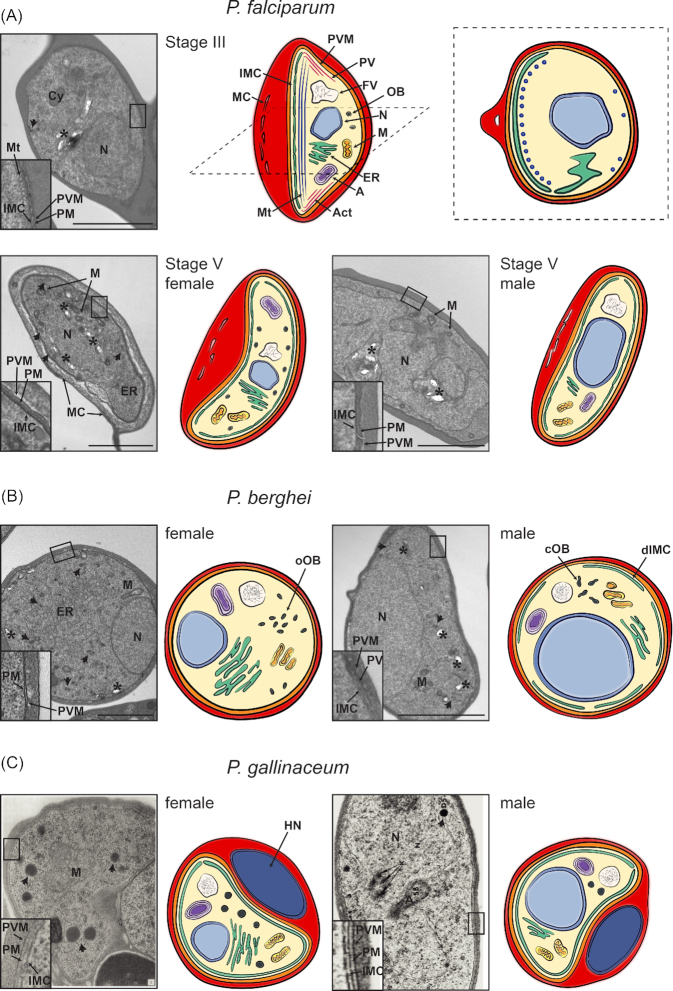
Morphological features of gametocytes across Plasmodium lineages. Shown are representative electron microscopy images and drawings of gametocytes of (A) human (P. falciparum), (B) rodent (P. berghei) and (C) avian (P. gallinaceum) malaria parasites. Mitochondria (M), apicoplast (A), nucleus (N), endoplasmatic reticulum (ER) and the food vacuole (FV) containing hemozoin crystals (marked with an asterisk) are found in all Plasmodium species in both male and female gametocytes. The gametocyte plasma membrane (PM) is in close association with the parasitophorous vacuolar membrane (PVM), and in most cases with the IMC (depicted in the closeup insets of the electron micrographs). Whilst the nucleus is smaller and more compact in female compared to male gametocytes, the ER is more developed in female gametocytes, in line with increased translation. Osmiophilic bodies (OB, marked with an arrow) are found in all species, with a higher number in female than in male gametocytes. (A) Ultrastructure of immature stage III and mature stage V P. falciparum gametocytes. Upper panel: Electron micrograph (left) and drawing (middle: longitudinal section, right: cross-section) of a stage III gametocyte. The IMC develops along one side of the gametocyte, supported by underlying microtubules (Mt)(depicted in the inset). Some microtubules are also found on the opposing side (see cross-section). Additionally, actin filaments (Act) are found, in particular at the tips of the developing gametocyte. MC are found within the erythrocyte cytoplasm. Lower panel: Electron microscope image and drawing of a female (left) and male (right) stage V gametocyte. The IMC plates now completely surround the gametocytes whilst actin and microtubules are absent. In the female gametocyte osmiophilic bodies are located along the periphery of the cell. In contrast they have not been described in male P. falciparum gametocytes. (B) Electron micrograph and drawing of mature female (left) and male (right) P. berghei gametocytes. Osmiophilic bodies are oval (oOB) and more abundant in female gametocytes, whilst male gametocytes have fewer and club-shaped osmiophilic bodies (cOB). The IMC is difficult to observe in female gametocytes and possibly absent, whilst a discontinuous IMC (dIMC) is clearly detectable in male P. berghei gametocytes (see insets) (Mons 1986). (C) Electron micrograph and scheme of mature female (left) and male (right) P. gallinaceum gametocytes. Electron micrographs are taken from (Aikawa et al. 1969)(female) and (Sterling and Aikawa 1973)(male) [PERMISSION PENDING]. In contrast to mammalian mature RBCs, avian mature RBCs are nucleated (depicted as host nucleus (HN)).

Comparative genomic view of gametocytogenesis across Plasmodium lineages. Heatmap showing the number of orthologous genes annotated as expressed specifically in P. falciparum gametocytes as defined by Pelle et al. (2015)(in rows) in major Plasmodium lineages. Apart from P. falciparum we have included P. vivax, P. berghei and P. gallinaceum. The orthologues of the P. falciparum gametocyte genes were retrieved from PlasmoDB (Aurrecoechea et al. 2009). The genes (rows) were reordered by hierarchical clustering using the R function hclust (www.R-project.org ). Using P. falciparum as a template (as there are no other species-specific sets of gametocyte genes) the majority of gametocyte-specific genes is conserved across the 4 species (see also Supplementary Table S1 (Supporting Information)). Most P. falciparum-specific genes in the gametocyte ring encode exported proteins. Interestingly, a putative zinc finger protein (PF3D7_1 134 600) has been lost in the rodent lineage only. Only a few immature and mature gametocyte genes are unique to P. falciparum, and most of them encode hypotheticals. Amongst the few exceptions are two putative protein kinases that have been selectively lost in primate (PF3D7_031 1400) and rodent malaria parasites (PF3D7_020 3100), respectively.
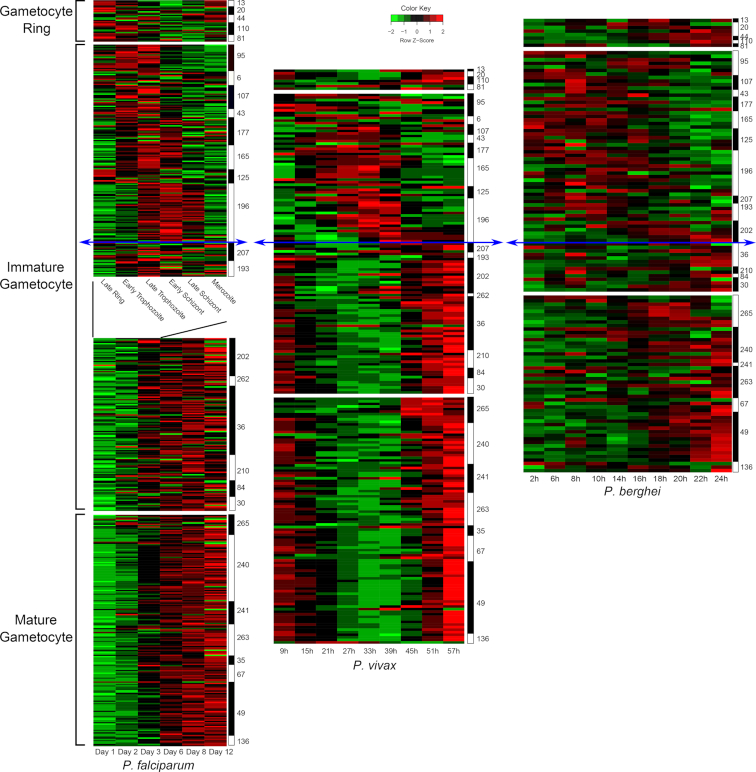
Transcriptional profiles across gametocyte development and Plasmodium lineages. Time-course data (Log2 Fold change) of gametocyte-specific genes were extracted from previously published datasets. For P. falciparum data are used from an asexual time course (Le Roch et al. 2003)(top part, in hours) and a gametocyte time course (Young et al.2005)(bottom part, in days). For P. vivax, data are used from an ex vivo time course (Bozdech et al. 2008)(in hours), and for P. berghei data are from an in vivo time course (Hoo et al. 2016)(in hours). Comparing the temporal transcription of the gametocyte-specific genes as defined in Pelle et al. and their orthologues in P. berghei and P. vivax shows that their profile is qualitatively conserved. The initial phase of development from the gametocyte ring stage to early immature gametocyte development shows a similar cascade of gene expression and takes a similar amount of time across the three species (20–39 hours). In contrast further gametocyte maturation lasts significantly longer in P. falciparum (9–10 days) than in P. vivax (about 17–20 hours) and P. berghei (about 4 hours). These differences in maturation time are likely related to the physiological differences in their respective hosts. The numbers on the right side indicate the cluster number as defined previously (Pelle et al. 2015). The blue arrows separate the time courses between the initial stages of development (top) with conserved duration and the later stages with variable in time scale.
References
-
- Aikawa M, Huff CG, Sprinz H. Comparative fine structure study of the gametocytes of avian, reptilian, and mammalian malarial parasites. J Ultrastruct Res. 1969;26:316–31. - PubMed
-
- Alano P, Roca L, Smith Det al. .. Plasmodium falciparum: parasites defective in early stages of gametocytogenesis. Exp Parasitol. 1995;81:227–35. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
