

New insights into cytoskeletal remodeling during platelet production
- PMID: 31220402
- PMCID: PMC6760864
- DOI: 10.1111/jth.14544
New insights into cytoskeletal remodeling during platelet production
Abstract
The past decade has brought unprecedented advances in our understanding of megakaryocyte (MK) biology and platelet production, processes that are strongly dependent on the cytoskeleton. Facilitated by technological innovations, such as new high-resolution imaging techniques (in vitro and in vivo) and lineage-specific gene knockout and reporter mouse strains, we are now able to visualize and characterize the molecular machinery required for MK development and proplatelet formation in live mice. Whole genome and RNA sequencing analysis of patients with rare platelet disorders, combined with targeted genetic interventions in mice, has led to the identification and characterization of numerous new genes important for MK development. Many of the genes important for proplatelet formation code for proteins that control cytoskeletal dynamics in cells, such as Rho GTPases and their downstream targets. In this review, we discuss how the final stages of MK development are controlled by the cellular cytoskeletons, and we compare changes in MK biology observed in patients and mice with mutations in cytoskeleton regulatory genes.
Keywords: bone marrow; cytoskeleton; megakaryocytes; platelets; proplatelet formation.
© 2019 International Society on Thrombosis and Haemostasis.
Conflict of interest statement
Conflicts of interest
The authors have no conflicts of interest to declare.
Figures
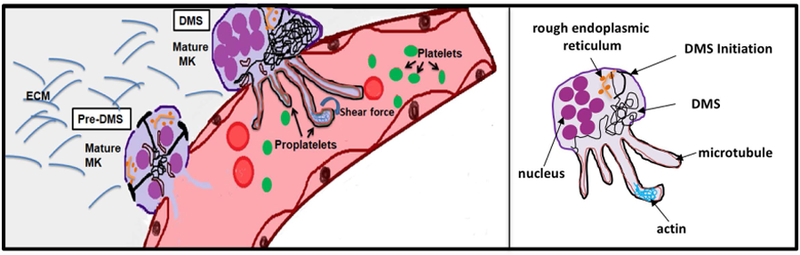
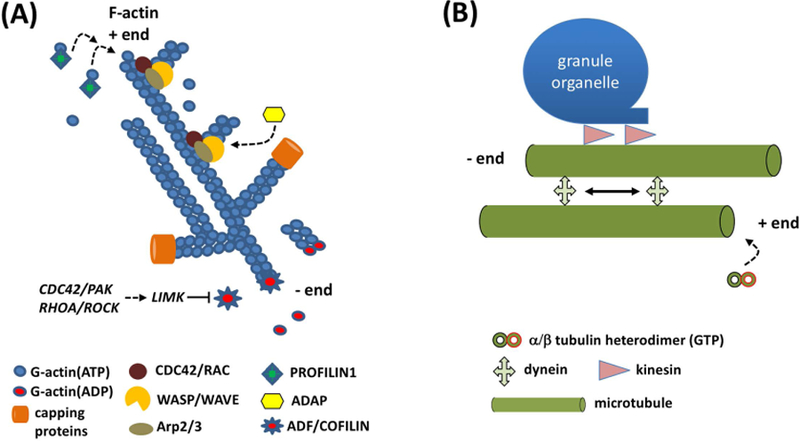
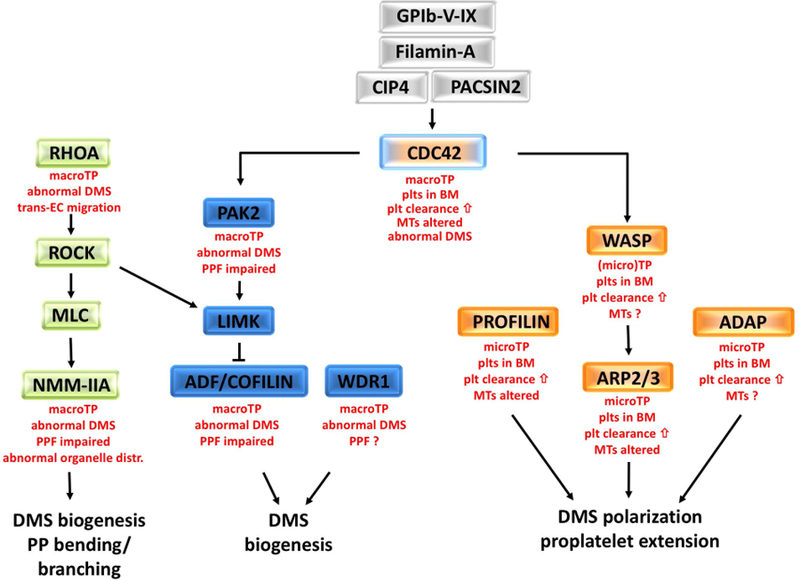
References
-
- Kaufman RM, Airo R, Pollack S, Crosby WH. Circulating megakaryocytes and platelet release in the lung. Blood 1965; 26(6): 720–31. - PubMed
-
- Lefrançais E, Ortiz-Muñoz G, Caudrillier A, Mallavia B, Liu F, Sayah DM, Thornton EE, Headley MB, David T, Coughlin SR, Krummel MF, Leavitt AD, Passegué E, Looney MR. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature 2017; 544(7648): 105–9. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
