

Adiponectin improves long-term potentiation in the 5XFAD mouse brain
- PMID: 31222110
- PMCID: PMC6586823
- DOI: 10.1038/s41598-019-45509-0
Adiponectin improves long-term potentiation in the 5XFAD mouse brain
Abstract
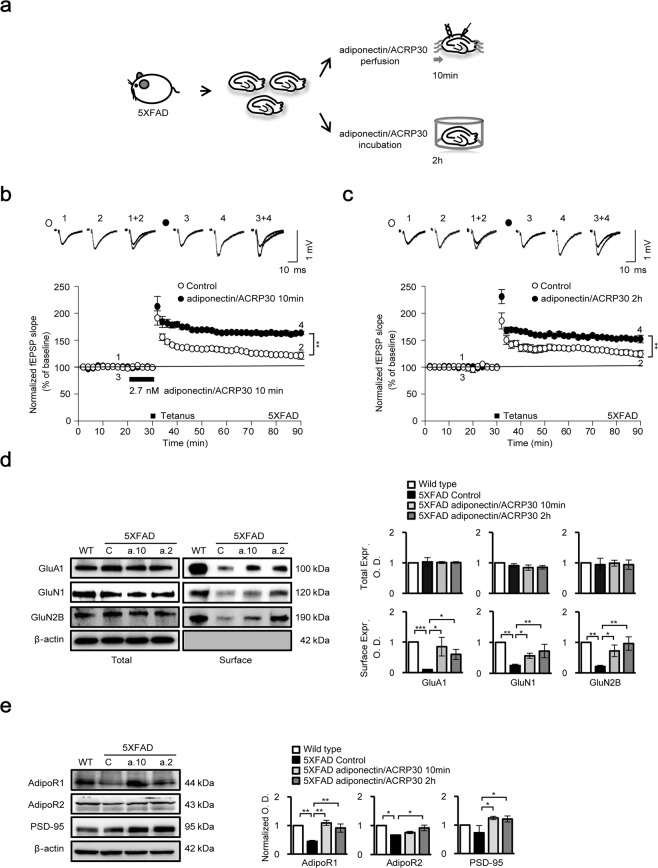
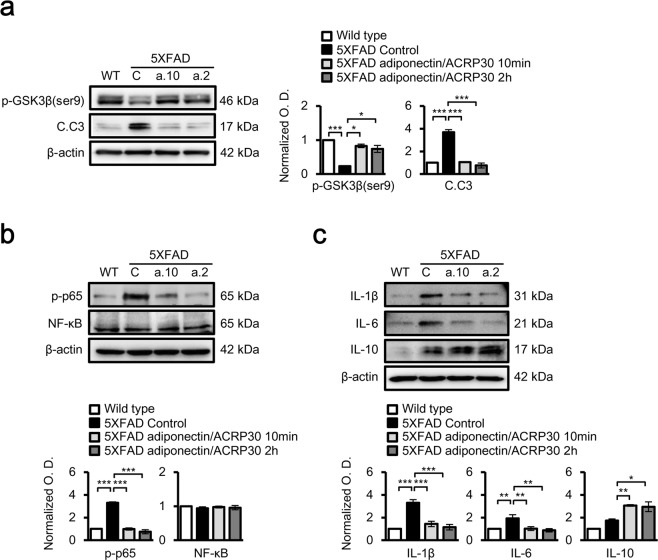
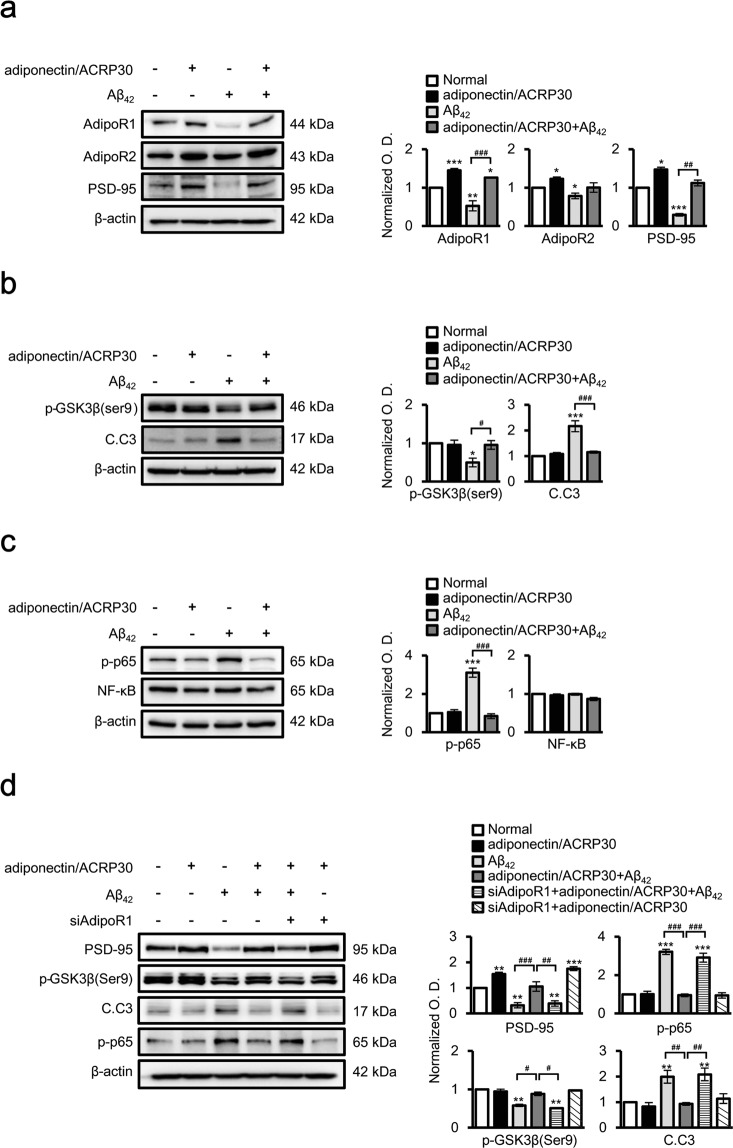
Adiponectin is an adipokine that regulates apoptosis, glucose and lipid metabolism, and insulin sensitivity in metabolic diseases. As recent studies have associated changes in adipokines and other metabolites in the central nervous system with a risk for Alzheimer's disease (AD), we investigated the effects of adiponectin treatment on hippocampal cells in the 5XFAD mouse model of AD and neuronal SH-SY5Y cells under amyloid beta toxicity. Adiponectin treatment reduced levels of cleaved caspase 3 and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) apoptosis signalling and decreased glycogen synthase kinase 3 beta (GSK3β) activation. Moreover, adiponectin treatment triggered long-term potentiation in the hippocampi of 5XFAD mice, which was associated with reduced expression of N-methyl-D-aspartate and its receptor as well as surface expression of the α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor. These findings suggest that adiponectin inhibits neuronal apoptosis and inflammatory mechanisms and promotes hippocampal long-term potentiation. Thus, adiponectin exhibits beneficial effect on hippocampal synaptic plasticity in Alzheimer's disease mouse model.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
β-amyloid inhibits hippocampal LTP through TNFR/IKK/NF-κB pathway.Neurol Res. 2018 Apr;40(4):268-276. doi: 10.1080/01616412.2018.1436872. Epub 2018 Feb 20. Neurol Res. 2018. PMID: 29458298
-
Oat Extract Avenanthramide-C Reverses Hippocampal Long-Term Potentiation Decline in Tg2576 Mice.Molecules. 2021 Oct 10;26(20):6105. doi: 10.3390/molecules26206105. Molecules. 2021. PMID: 34684684 Free PMC article.
-
Adiponectin controls the apoptosis and the expression of tight junction proteins in brain endothelial cells through AdipoR1 under beta amyloid toxicity.Cell Death Dis. 2017 Oct 12;8(10):e3102. doi: 10.1038/cddis.2017.491. Cell Death Dis. 2017. PMID: 29022894 Free PMC article.
-
Plasticity, hippocampal place cells, and cognitive maps.Arch Neurol. 2001 Jun;58(6):874-81. doi: 10.1001/archneur.58.6.874. Arch Neurol. 2001. PMID: 11405801 Review.
-
Impaired autophagy and APP processing in Alzheimer's disease: The potential role of Beclin 1 interactome.Prog Neurobiol. 2013 Jul-Aug;106-107:33-54. doi: 10.1016/j.pneurobio.2013.06.002. Epub 2013 Jul 1. Prog Neurobiol. 2013. PMID: 23827971 Review.
Cited by
-
Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats.Cells. 2024 Jan 17;13(2):182. doi: 10.3390/cells13020182. Cells. 2024. PMID: 38247873 Free PMC article.
-
The Novel Perspectives of Adipokines on Brain Health.Int J Mol Sci. 2019 Nov 11;20(22):5638. doi: 10.3390/ijms20225638. Int J Mol Sci. 2019. PMID: 31718027 Free PMC article. Review.
-
Chronic AdipoRon Treatment Mimics the Effects of Physical Exercise on Restoring Hippocampal Neuroplasticity in Diabetic Mice.Mol Neurobiol. 2021 Sep;58(9):4666-4681. doi: 10.1007/s12035-021-02441-7. Epub 2021 Jun 23. Mol Neurobiol. 2021. PMID: 34164760 Free PMC article.
-
Adiponectin Mediates the Protection of H2S Against Chronic Restraint Stress-Induced Cognitive Impairment via Attenuating Hippocampal Damage.Front Behav Neurosci. 2021 May 5;15:623644. doi: 10.3389/fnbeh.2021.623644. eCollection 2021. Front Behav Neurosci. 2021. PMID: 34025367 Free PMC article.
-
Central Adiponectin Signaling - A Metabolic Regulator in Support of Brain Plasticity.Brain Plast. 2022 Oct 21;8(1):79-96. doi: 10.3233/BPL-220138. eCollection 2022. Brain Plast. 2022. PMID: 36448043 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
