

Mechanisms and biomedical implications of -1 programmed ribosome frameshifting on viral and bacterial mRNAs
- PMID: 31222875
- PMCID: PMC6771820
- DOI: 10.1002/1873-3468.13478
Mechanisms and biomedical implications of -1 programmed ribosome frameshifting on viral and bacterial mRNAs
Abstract
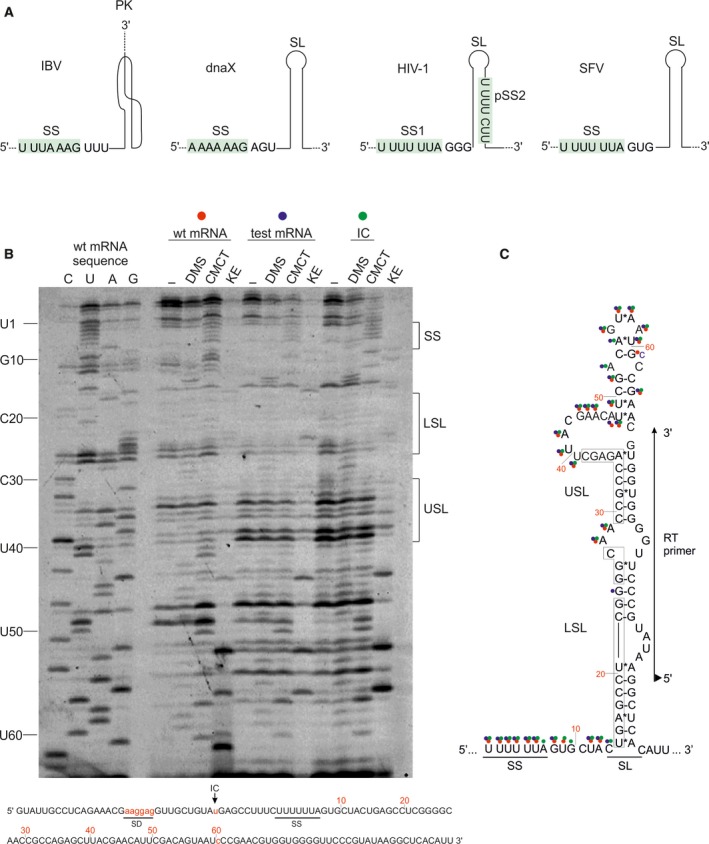
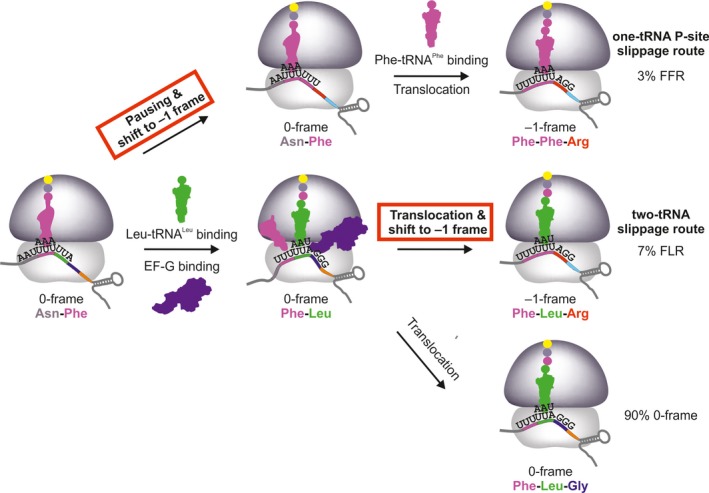
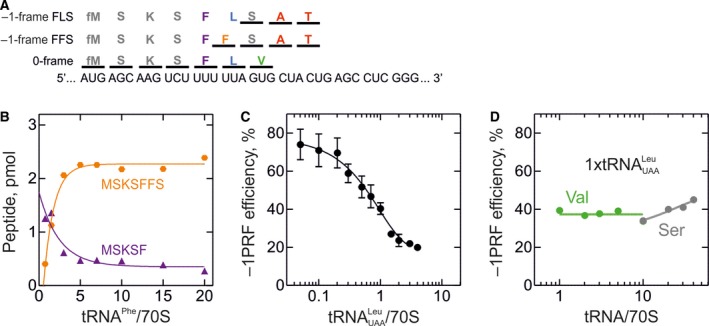
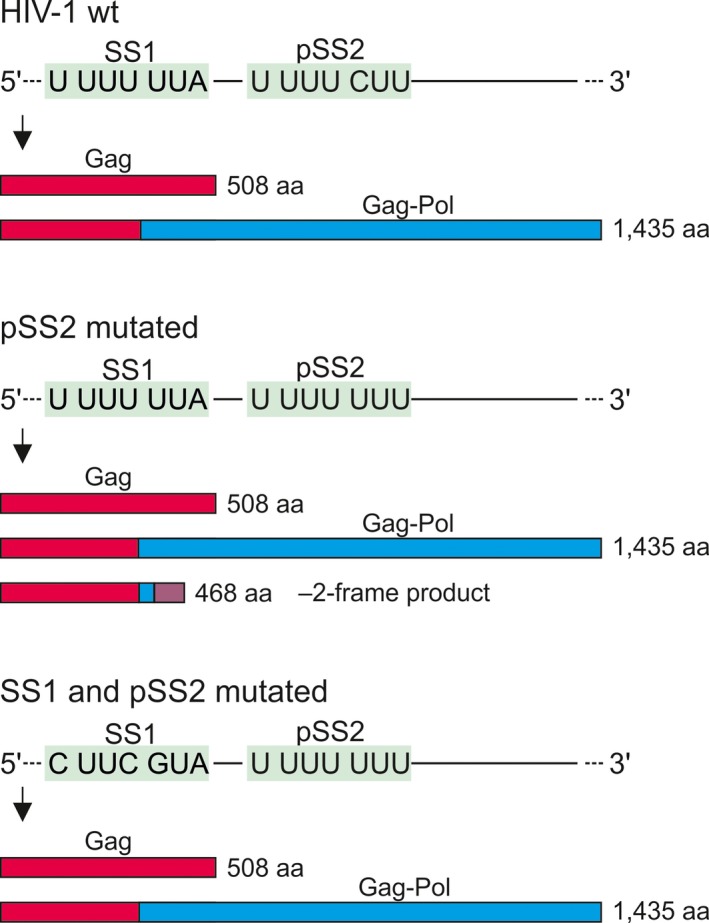
Some proteins are expressed as a result of a ribosome frameshifting event that is facilitated by a slippery site and downstream secondary structure elements in the mRNA. This review summarizes recent progress in understanding mechanisms of -1 frameshifting in several viral genes, including IBV 1a/1b, HIV-1 gag-pol, and SFV 6K, and in Escherichia coli dnaX. The exact frameshifting route depends on the availability of aminoacyl-tRNAs: the ribosome normally slips into the -1-frame during tRNA translocation, but can also frameshift during decoding at condition when aminoacyl-tRNA is in limited supply. Different frameshifting routes and additional slippery sites allow viruses to maintain a constant production of their key proteins. The emerging idea that tRNA pools are important for frameshifting provides new direction for developing antiviral therapies.
Keywords: RNA; tRNA; frameshifting; protein synthesis; recoding; regulation; ribosome; translation.
© 2019 The Authors. FEBS Letters published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Figures
References
-
- Demeshkina N, Jenner L, Westhof E, Yusupov M and Yusupova G (2013) New structural insights into the decoding mechanism: translation infidelity via a G.U pair with Watson‐Crick geometry. FEBS Lett 587, 1848–1857. - PubMed
-
- Rodnina MV (2012) Quality control of mRNA decoding on the bacterial ribosome. Adv Protein Chem Struct Biol 86, 95–128. - PubMed
-
- Ramakrishnan V (2014) The ribosome emerges from a black box. Cell 159, 979–984. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
