

Reciprocal regulation of TORC signaling and tRNA modifications by Elongator enforces nutrient-dependent cell fate
- PMID: 31223645
- PMCID: PMC6584457
- DOI: 10.1126/sciadv.aav0184
Reciprocal regulation of TORC signaling and tRNA modifications by Elongator enforces nutrient-dependent cell fate
Abstract
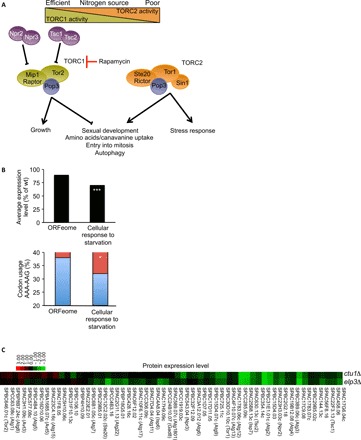
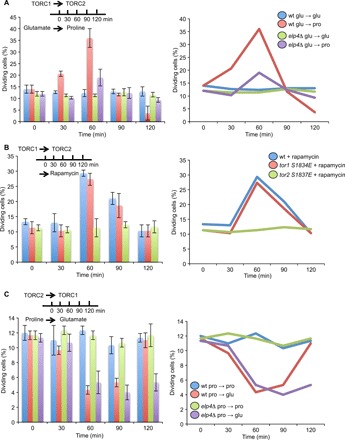
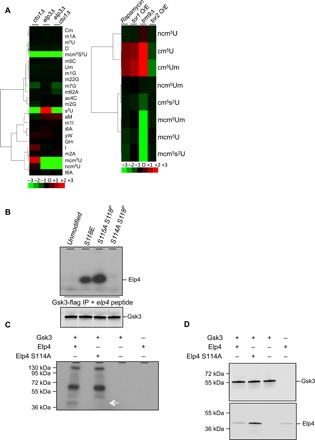
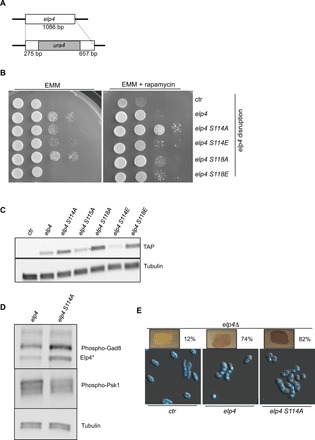
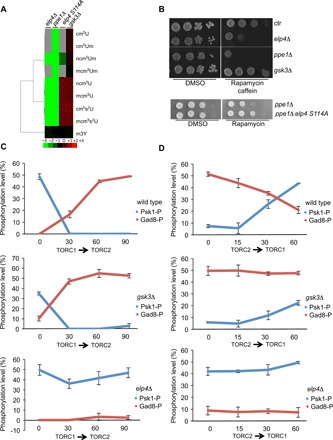
Nutrient availability has a profound impact on cell fate. Upon nitrogen starvation, wild-type fission yeast cells uncouple cell growth from cell division to generate small, round-shaped cells that are competent for sexual differentiation. The TORC1 (TOR complex 1) and TORC2 complexes exert opposite controls on cell growth and cell differentiation, but little is known about how their activity is coordinated. We show that transfer RNA (tRNA) modifications by Elongator are critical for this regulation by promoting the translation of both key components of TORC2 and repressors of TORC1. We further identified the TORC2 pathway as an activator of Elongator by down-regulating a Gsk3 (glycogen synthase kinase 3)-dependent inhibitory phosphorylation of Elongator. Therefore, a feedback control is operating between TOR complex (TORC) signaling and tRNA modification by Elongator to enforce the advancement of mitosis that precedes cell differentiation.
Figures

References
-
- McDonald I. J., Callejam G. B., Johnson B. F., Conjugation in chemostat cultures of Schizosaccharomyces pombe. J. Gen. Microbiol. 128, 1981–1987 (1982).
-
- Egel R., Commitment to meiosis in fission yeast. Mol. Gen. Genet. 121, 277–284 (1973).
-
- Kim D.-H., Sarbassov D. D., Ali S. M., King J. E., Latek R. R., Erdjument-Bromage H., Tempst P., Sabatini D. M., mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery. Cell 110, 163–175 (2002). - PubMed
-
- Montagne J., Stewart M. J., Stocker H., Hafen E., Kozma S. C., Thomas G., Drosophila S6 kinase: A regulator of cell size. Science 285, 2126–2129 (1999). - PubMed
-
- Álvarez B., Moreno S., Fission yeast Tor2 promotes cell growth and represses cell differentiation. J. Cell Sci. 119, 4475–4485 (2006). - PubMed
