

The emerging role of minor intron splicing in neurological disorders
- PMID: 31225466
- PMCID: PMC6558932
- DOI: 10.15698/cst2018.03.126
The emerging role of minor intron splicing in neurological disorders
Abstract
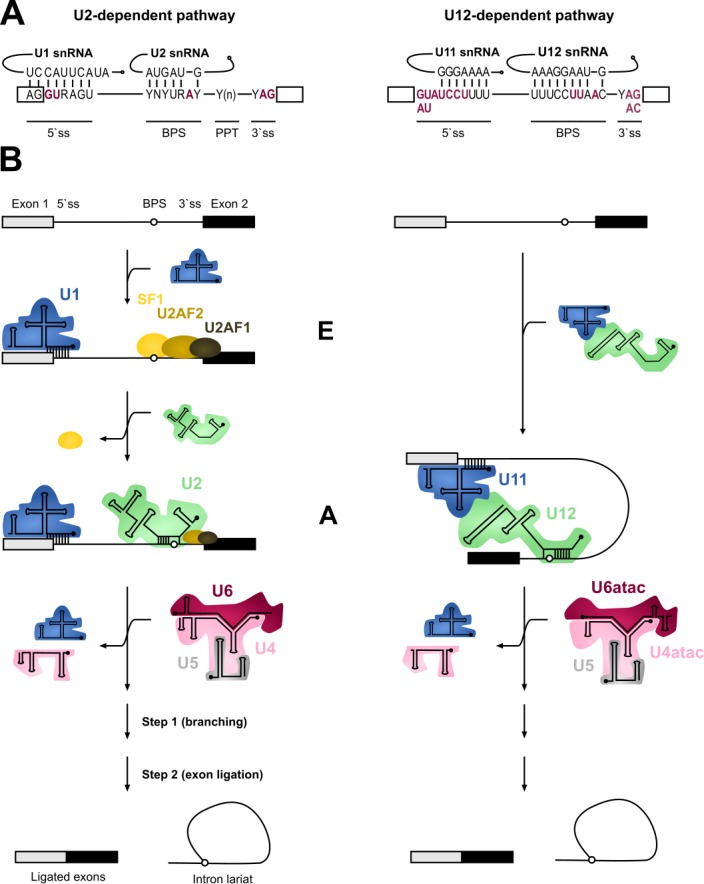
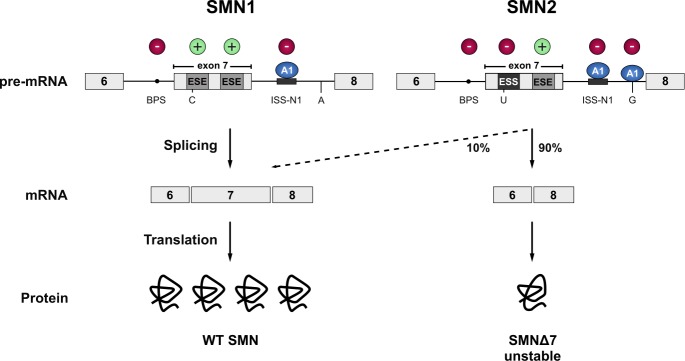
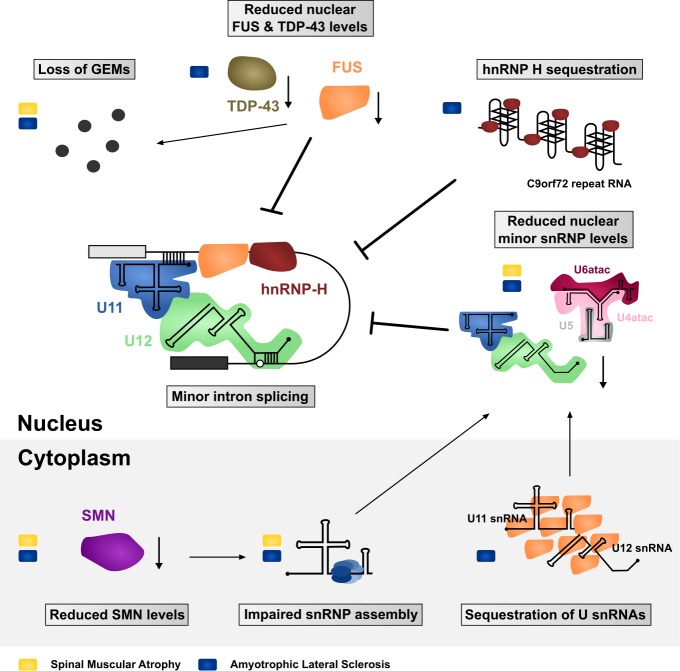
Pre-mRNA splicing is an essential step in eukaryotic gene expression. Mutations in cis-acting sequence elements within pre-mRNA molecules or trans-acting factors involved in pre-mRNA processing have both been linked to splicing dysfunction that give rise to a large number of human diseases. These mutations typically affect the major splicing pathway, which excises more than 99% of all introns in humans. However, approximately 700-800 human introns feature divergent intron consensus sequences at their 5' and 3' ends and are recognized by a separate pre-mRNA processing machinery denoted as the minor spliceosome. This spliceosome has been studied less than its major counterpart, but has received increasing attention during the last few years as a novel pathomechanistic player on the stage in neurodevelopmental and neurodegenerative diseases. Here, we review the current knowledge on minor spliceosome function and discuss its potential pathomechanistic role and impact in neurodegeneration.
Keywords: ALS; FUS; SMA; TDP-43; minor spliceosome; neurodegeneration; pre-mRNA splicing.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Miscellaneous