

Putting together and taking apart: assembly and disassembly of the Rad51 nucleoprotein filament in DNA repair and genome stability
- PMID: 31225474
- PMCID: PMC6551702
- DOI: 10.15698/cst2018.05.134
Putting together and taking apart: assembly and disassembly of the Rad51 nucleoprotein filament in DNA repair and genome stability
Abstract
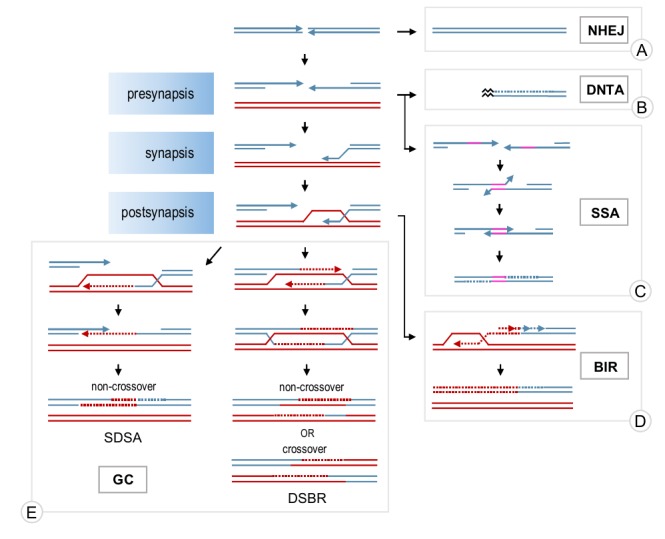
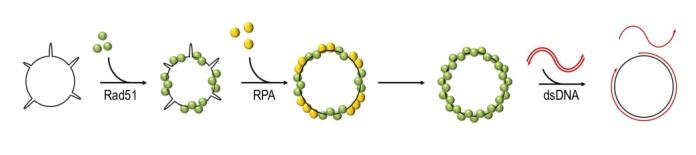
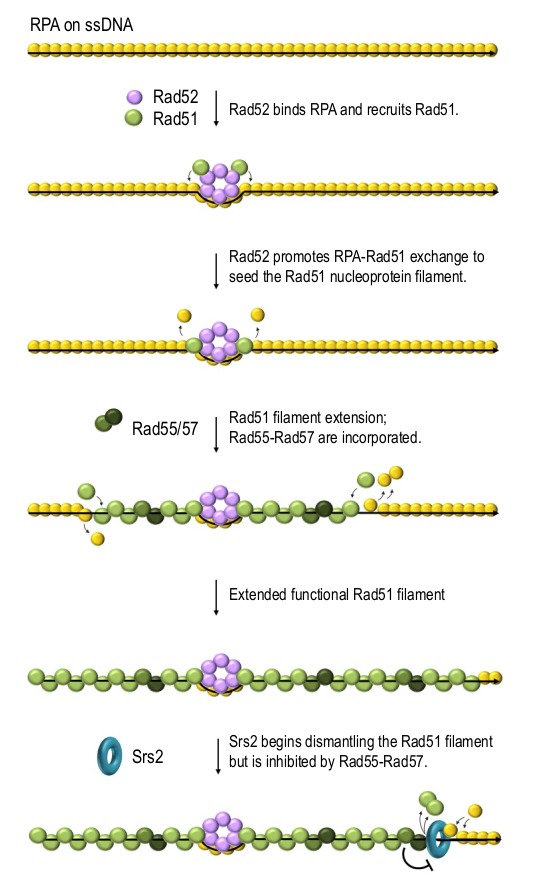
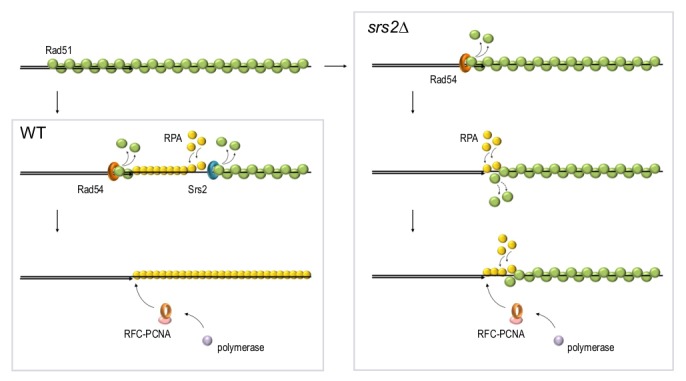
Homologous recombination is a key mechanism providing both genome stability and genetic diversity in all living organisms. Recombinases play a central role in this pathway: multiple protein subunits of Rad51 or its orthologues bind single-stranded DNA to form a nucleoprotein filament which is essential for initiating recombination events. Multiple factors are involved in the regulation of this step, both positively and negatively. In this review, we discuss Rad51 nucleoprotein assembly and disassembly, how it is regulated and what functional significance it has in genome maintenance.
Keywords: DNA repair; Rad51 filament; Rad51 regulation; double-stranded DNA break; homologous recombination.
Conflict of interest statement
Conflict of interest: The authors declare no conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Research Materials