

New pathologic mechanisms in nucleotide repeat expansion disorders
- PMID: 31229686
- PMCID: PMC6689435
- DOI: 10.1016/j.nbd.2019.104515
New pathologic mechanisms in nucleotide repeat expansion disorders
Abstract
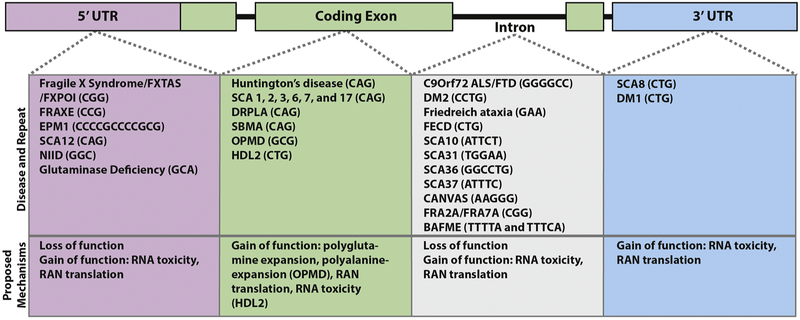
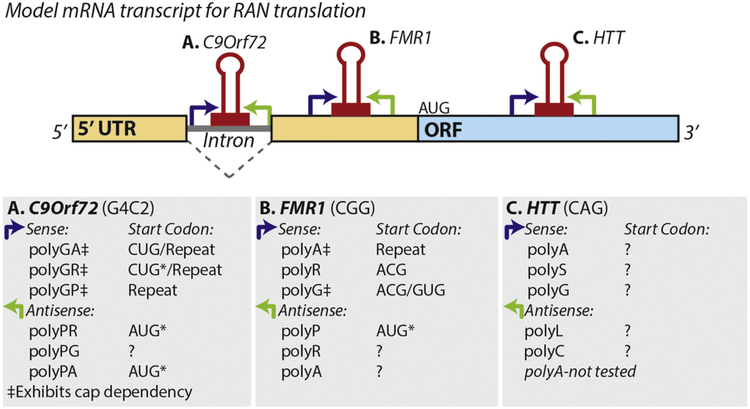
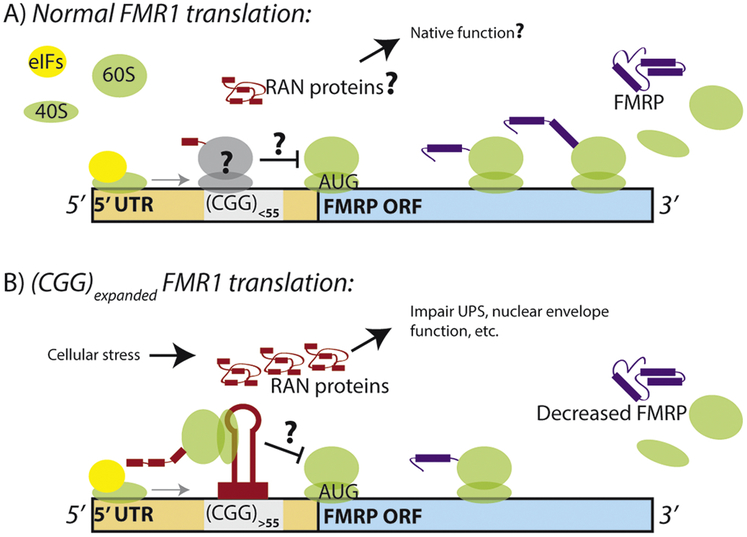
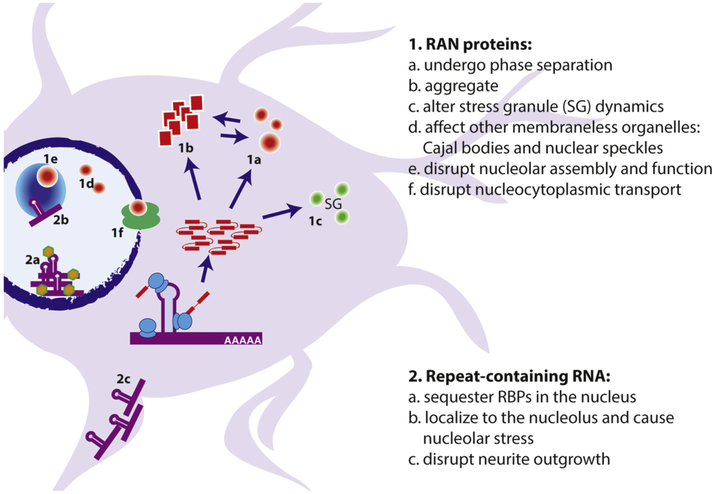
Tandem microsatellite repeats are common throughout the human genome and intrinsically unstable, exhibiting expansions and contractions both somatically and across generations. Instability in a small subset of these repeats are currently linked to human disease, although recent findings suggest more disease-causing repeats await discovery. These nucleotide repeat expansion disorders (NREDs) primarily affect the nervous system and commonly lead to neurodegeneration through toxic protein gain-of-function, protein loss-of-function, and toxic RNA gain-of-function mechanisms. However, the lines between these categories have blurred with recent findings of unconventional Repeat Associated Non-AUG (RAN) translation from putatively non-coding regions of the genome. Here we review two emerging topics in NREDs: 1) The mechanisms by which RAN translation occurs and its role in disease pathogenesis and 2) How nucleotide repeats as RNA and translated proteins influence liquid-liquid phase separation, membraneless organelle dynamics, and nucleocytoplasmic transport. We examine these topics with a particular eye on two repeats: the CGG repeat expansion responsible for Fragile X syndrome and Fragile X-associated Tremor Ataxia Syndrome (FXTAS) and the intronic GGGGCC repeat expansion in C9orf72, the most common inherited cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Our thesis is that these emerging disease mechanisms can inform a broader understanding of the native roles of microsatellites in cellular function and that aberrations in these native processes provide clues to novel therapeutic strategies for these currently untreatable disorders.
Published by Elsevier Inc.
Figures
References
-
- Anderson P, and Kedersha N. 2009. 'RNA granules: post-transcriptional and epigenetic modulators of gene expression', Nat Rev Mol Cell Biol, 10: 430–6. - PubMed
-
- Andrew SE, Goldberg YP, Kremer B, Telenius H, Theilmann J, Adam S, Starr E, Squitieri F, Lin B, Kalchman MA, and et al. 1993. 'The relationship between trinucleotide (CAG) repeat length and clinical features of Huntington's disease', Nat Genet, 4: 398–403. - PubMed
-
- Arocena DG, Iwahashi CK, Won N, Beilina A, Ludwig AL, Tassone F, Schwartz PH, and Hagerman PJ. 2005. 'Induction of inclusion formation and disruption of lamin A/C structure by premutation CGG-repeat RNA in human cultured neural cells', Hum Mol Genet, 14: 3661–71. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
