

Transforming the Choice Outcome to an Action Plan in Monkey Lateral Prefrontal Cortex: A Neural Circuit Model
- PMID: 31230761
- PMCID: PMC7370701
- DOI: 10.1016/j.neuron.2019.05.032
Transforming the Choice Outcome to an Action Plan in Monkey Lateral Prefrontal Cortex: A Neural Circuit Model
Abstract
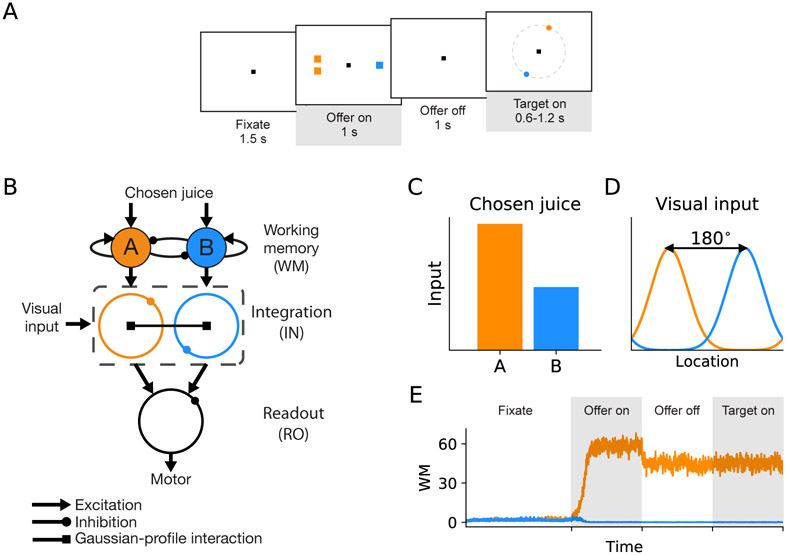
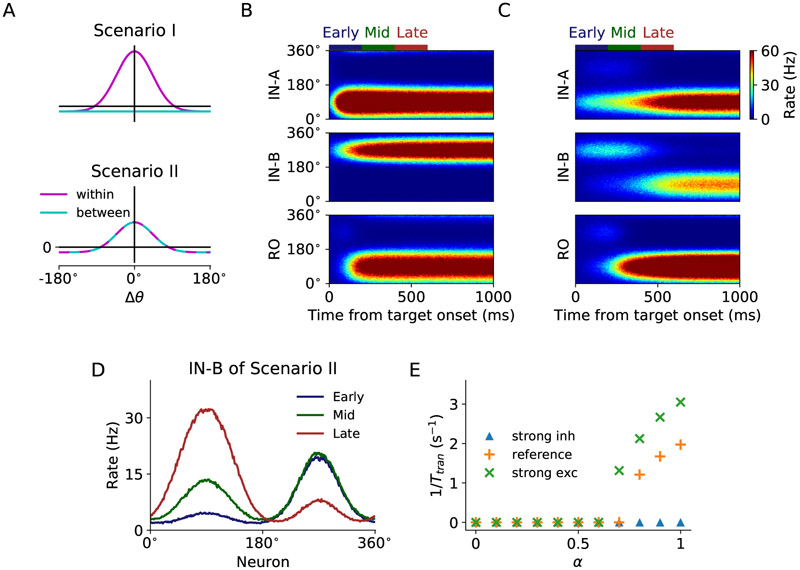
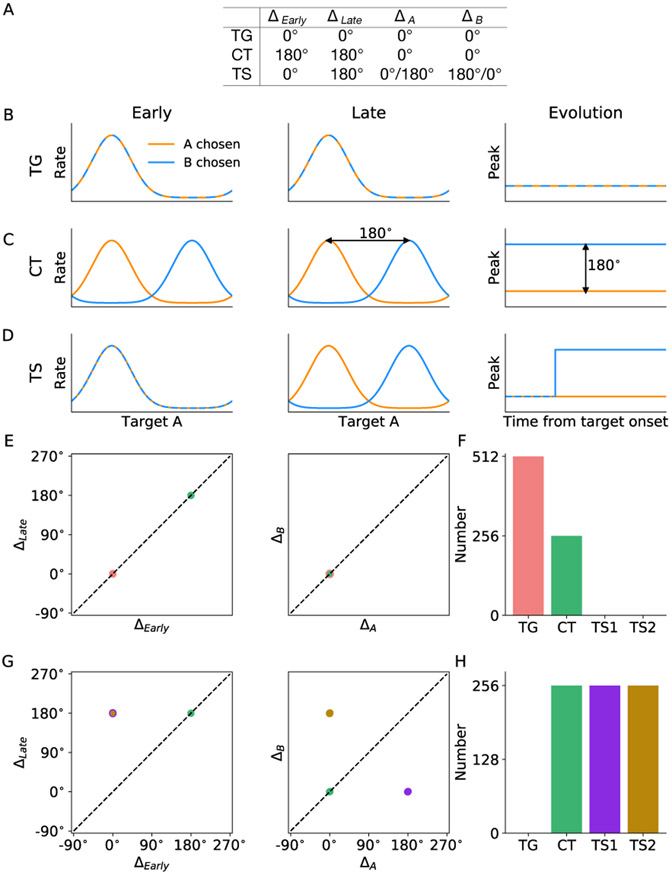
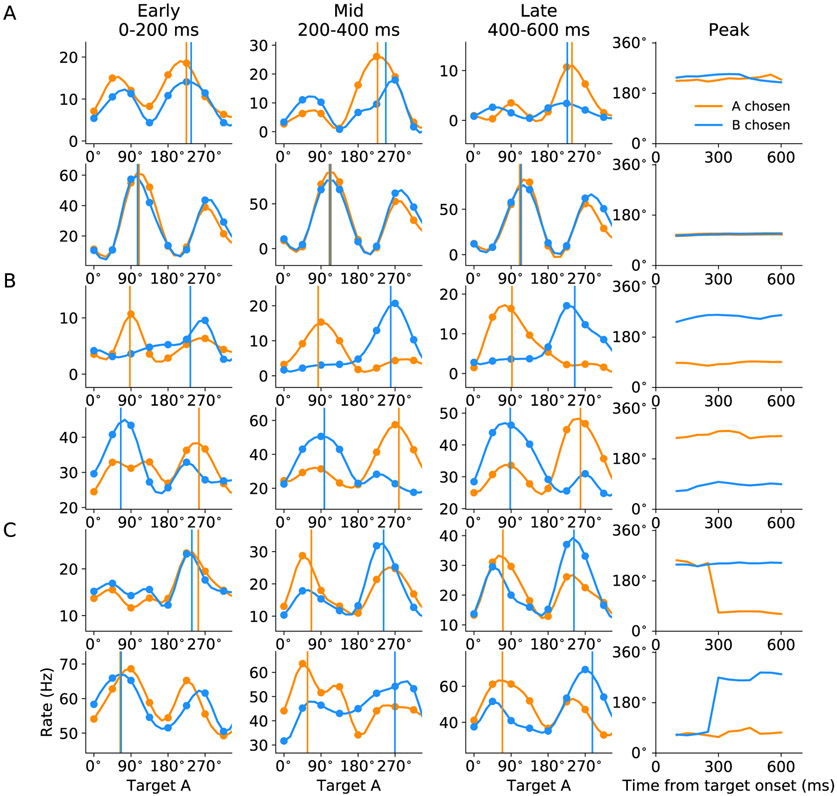
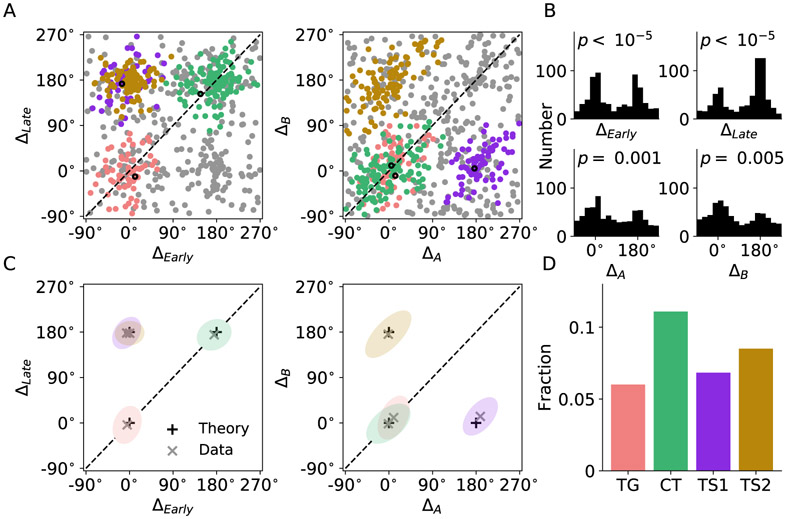
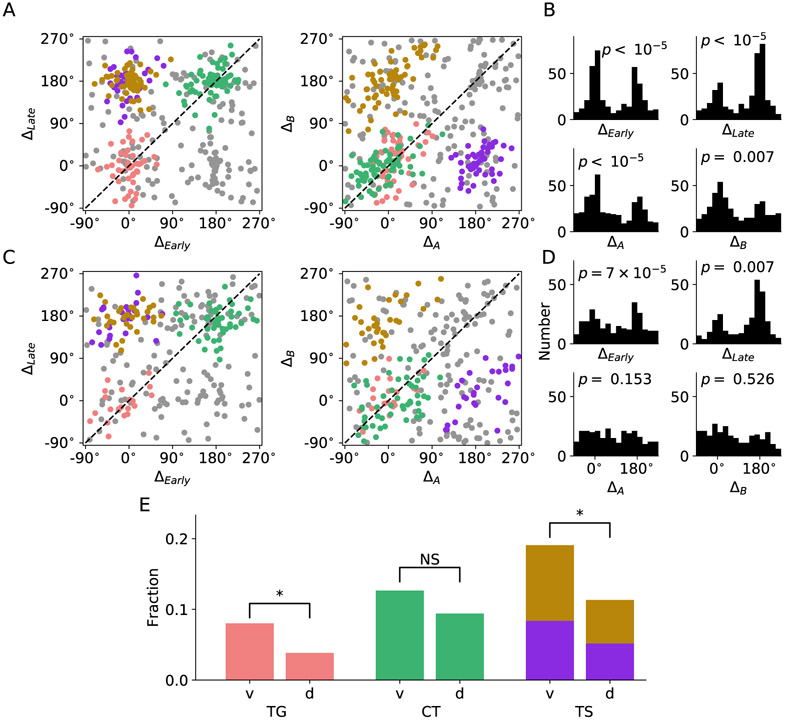
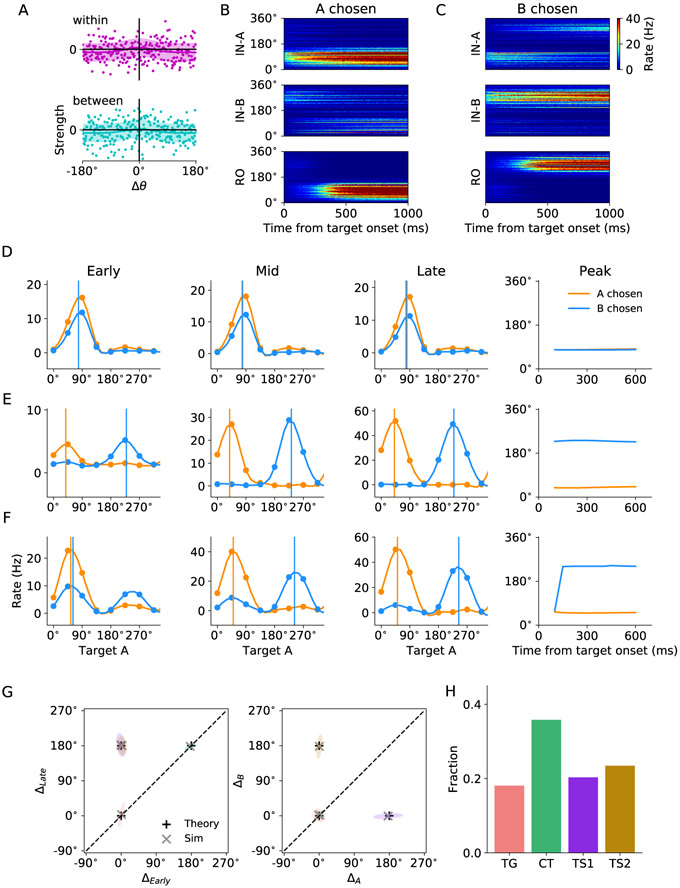
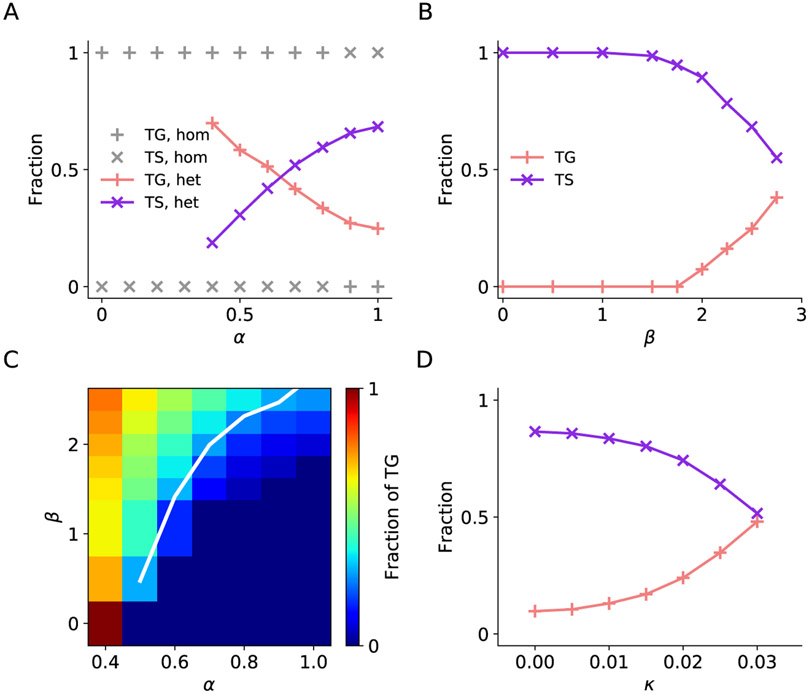
In economic decisions, we make a good-based choice first, then we transform the outcome into an action to obtain the good. To elucidate the network mechanisms for such transformation, we constructed a neural circuit model consisting of modules representing choice, integration of choice with target locations, and the final action plan. We examined three scenarios regarding how the final action plan could emerge in the neural circuit and compared their implications with experimental data. Our model with heterogeneous connectivity predicts the coexistence of three types of neurons with distinct functions, confirmed by analyzing the neural activity in the lateral prefrontal cortex (LPFC) of behaving monkeys. We obtained a much more distinct classification of functional neuron types in the ventral than the dorsal region of LPFC, suggesting that the action plan is initially generated in ventral LPFC. Our model offers a biologically plausible neural circuit architecture that implements good-to-action transformation during economic choice.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Contributions of orbitofrontal and lateral prefrontal cortices to economic choice and the good-to-action transformation.Neuron. 2014 Mar 5;81(5):1140-1151. doi: 10.1016/j.neuron.2014.01.008. Epub 2014 Feb 13. Neuron. 2014. PMID: 24529981 Free PMC article.
-
Differential coding of goals and actions in ventral and dorsal corticostriatal circuits during goal-directed behavior.Cell Rep. 2022 Jan 4;38(1):110198. doi: 10.1016/j.celrep.2021.110198. Cell Rep. 2022. PMID: 34986350 Free PMC article.
-
Working Memory and Decision-Making in a Frontoparietal Circuit Model.J Neurosci. 2017 Dec 13;37(50):12167-12186. doi: 10.1523/JNEUROSCI.0343-17.2017. Epub 2017 Nov 7. J Neurosci. 2017. PMID: 29114071 Free PMC article.
-
[Neural mechanisms of decision making].Brain Nerve. 2008 Sep;60(9):1017-27. Brain Nerve. 2008. PMID: 18807936 Review. Japanese.
-
Reward-dependent learning in neuronal networks for planning and decision making.Prog Brain Res. 2000;126:217-29. doi: 10.1016/S0079-6123(00)26016-0. Prog Brain Res. 2000. PMID: 11105649 Review.
Cited by
-
A structural and functional subdivision in central orbitofrontal cortex.Nat Commun. 2022 Jun 24;13(1):3623. doi: 10.1038/s41467-022-31273-9. Nat Commun. 2022. PMID: 35750659 Free PMC article.
-
Category learning in a recurrent neural network with reinforcement learning.Front Psychiatry. 2022 Oct 25;13:1008011. doi: 10.3389/fpsyt.2022.1008011. eCollection 2022. Front Psychiatry. 2022. PMID: 36387007 Free PMC article.
-
Value dynamics affect choice preparation during decision-making.Nat Neurosci. 2023 Sep;26(9):1575-1583. doi: 10.1038/s41593-023-01407-3. Epub 2023 Aug 10. Nat Neurosci. 2023. PMID: 37563295 Free PMC article.
-
Taking stock of value in the orbitofrontal cortex.Nat Rev Neurosci. 2022 Jul;23(7):428-438. doi: 10.1038/s41583-022-00589-2. Epub 2022 Apr 25. Nat Rev Neurosci. 2022. PMID: 35468999 Free PMC article. Review.
-
Neuronal origins of reduced accuracy and biases in economic choices under sequential offers.Elife. 2022 Apr 13;11:e75910. doi: 10.7554/eLife.75910. Elife. 2022. PMID: 35416775 Free PMC article.
References
-
- Abbott L, and Chance FS (2005). Drivers and modulators from push-pull and balanced synaptic input. Prog. Brain Res 149, 147–155. - PubMed
-
- Berens P (2009). CircStat : a MATLAB toolbox for circular statistics. J. Stat. Softw 31, 1–21.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
