

The R941L mutation in MYH14 disrupts mitochondrial fission and associates with peripheral neuropathy
- PMID: 31231018
- PMCID: PMC6642256
- DOI: 10.1016/j.ebiom.2019.06.018
The R941L mutation in MYH14 disrupts mitochondrial fission and associates with peripheral neuropathy
Abstract
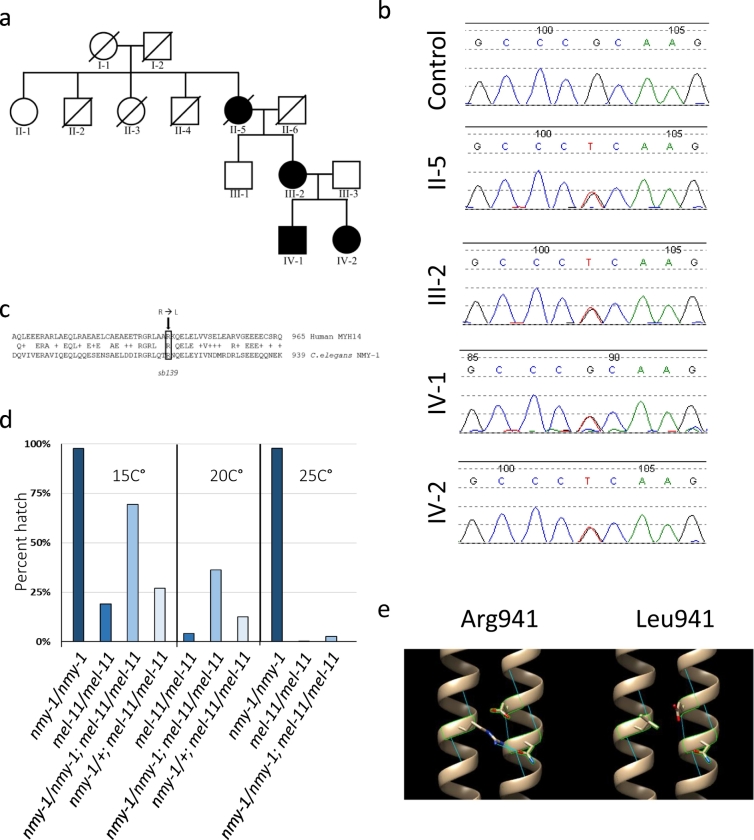
Background: Peripheral neuropathies are often caused by disruption of genes responsible for myelination or axonal transport. In particular, impairment in mitochondrial fission and fusion are known causes of peripheral neuropathies. However, the causal mechanisms for peripheral neuropathy gene mutations are not always known. While loss of function mutations in MYH14 typically cause non-syndromic hearing loss, the recently described R941L mutation in MYH14, encoding the non-muscle myosin protein isoform NMIIC, leads to a complex clinical presentation with an unexplained peripheral neuropathy phenotype.
Methods: Confocal microscopy was used to examine mitochondrial dynamics in MYH14 patient fibroblast cells, as well as U2OS and M17 cells overexpressing NMIIC. The consequence of the R941L mutation on myosin activity was modeled in C. elegans.
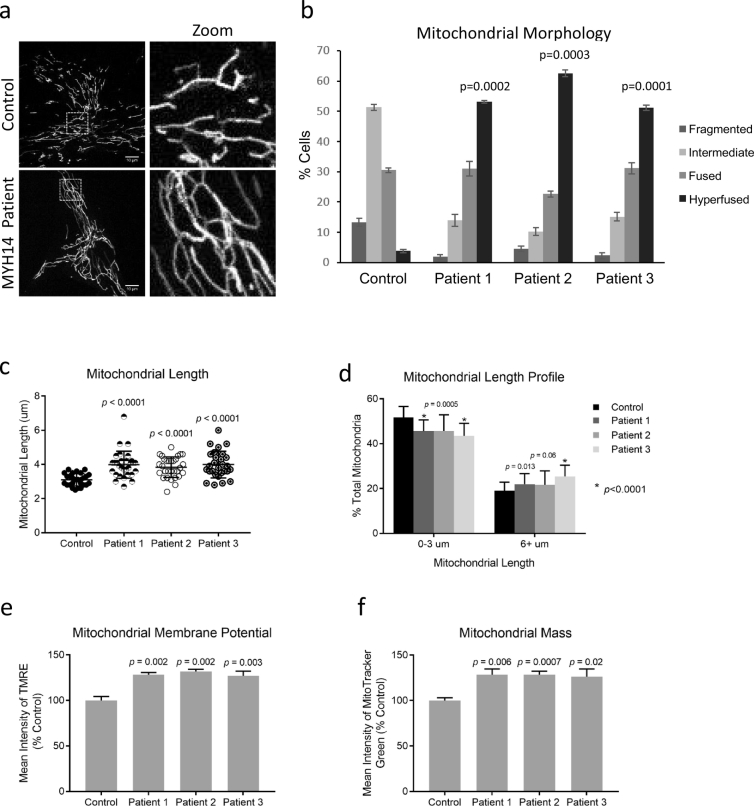
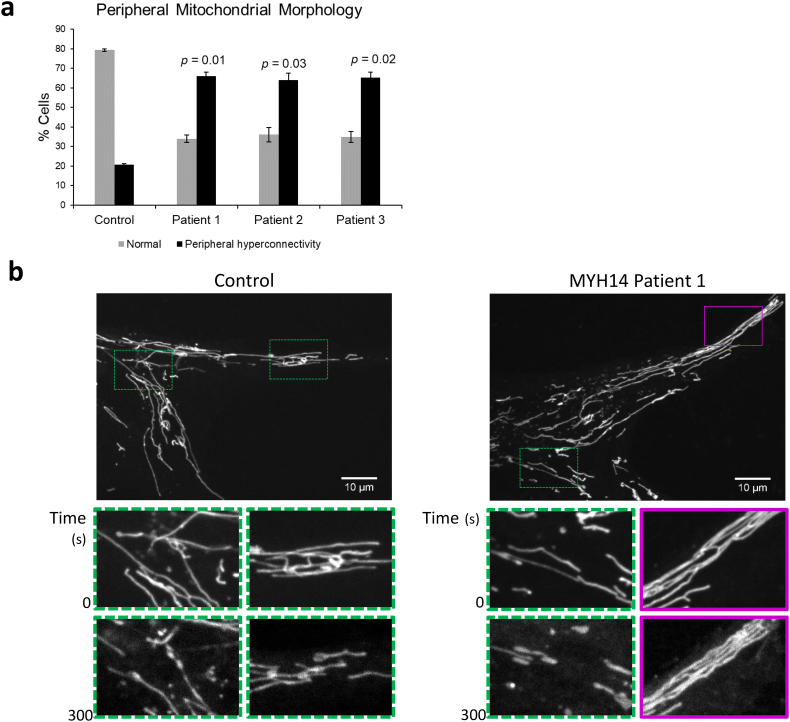
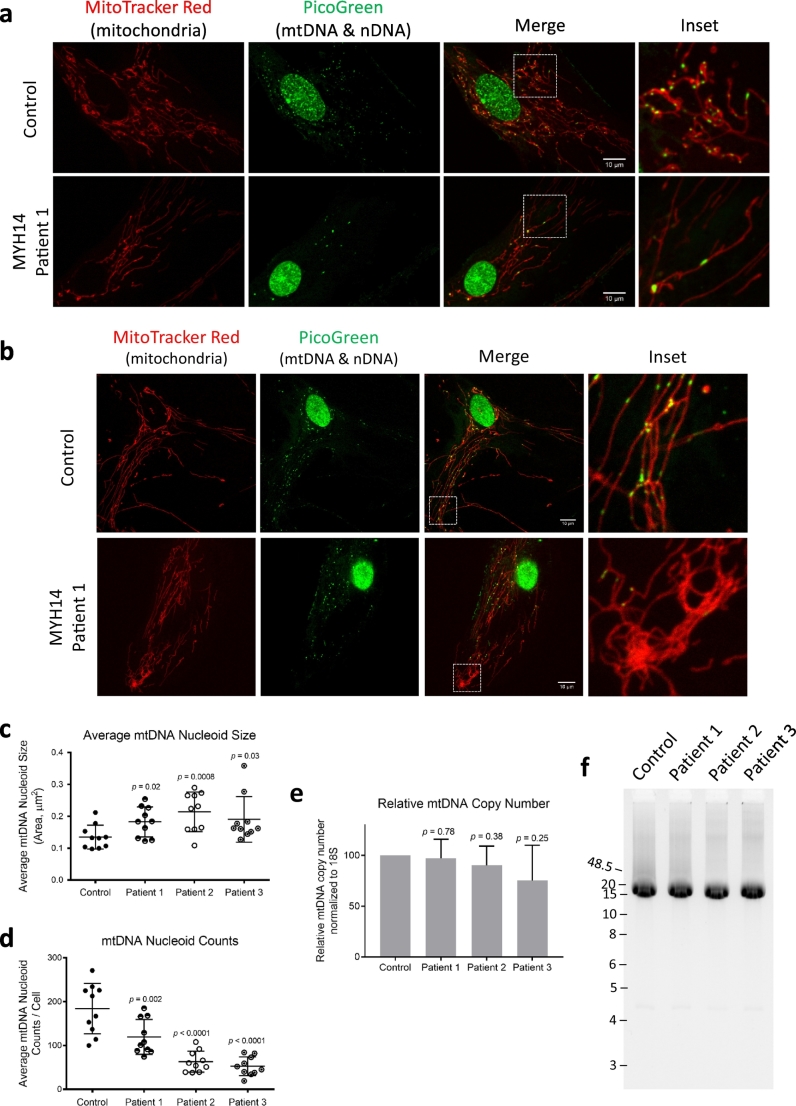
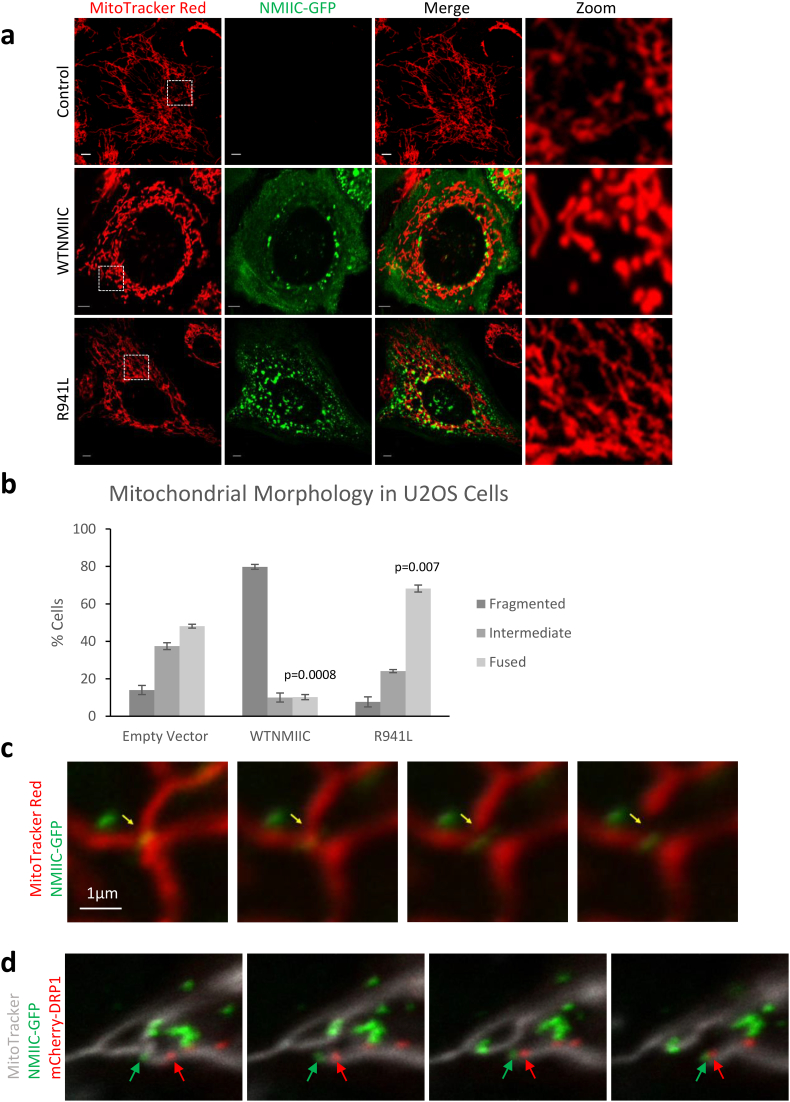
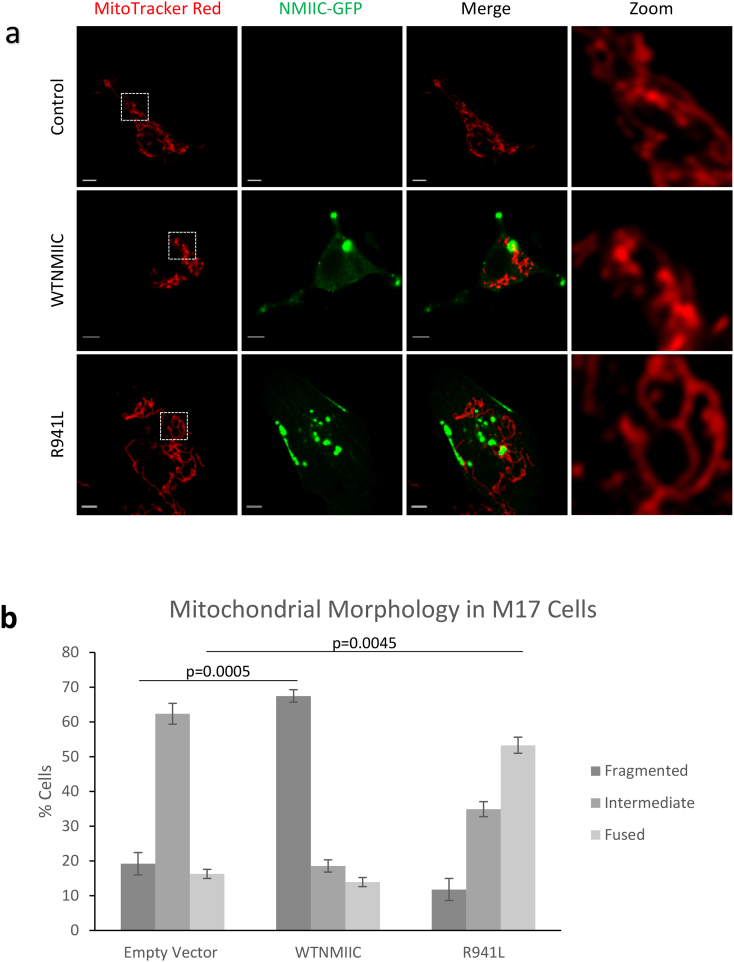
Findings: We describe the third family carrying the R941L mutation in MYH14, and demonstrate that the R941L mutation impairs non-muscle myosin protein function. To better understand the molecular basis of the peripheral neuropathy phenotype associated with the R941L mutation, which has been hindered by the fact that NMIIC is largely uncharacterized, we have established a previously unrecognized biological role for NMIIC in mediating mitochondrial fission in human cells. Notably, the R941L mutation acts in a dominant-negative fashion to inhibit mitochondrial fission, especially in the cell periphery. In addition, we observed alterations to the organization of the mitochondrial genome.
Interpretation: As impairments in mitochondrial fission cause peripheral neuropathy, this insight into the function of NMIIC likely explains the peripheral neuropathy phenotype associated with the R941L mutation. FUND: This study was supported by the Alberta Children's Hospital Research Institute, the Canadian Institutes of Health Research and the Care4Rare Canada Consortium.
Keywords: Caenorhabditis elegans; Mitochondria; Mitochondrial fission; Non-muscle myosin; Peripheral neuropathy; mtDNA.
Copyright © 2019. Published by Elsevier B.V.
Figures
Comment in
-
Neuropathy due to impaired axonal transport of non-fragmented mitochondria in MYH14 mutation carriers.EBioMedicine. 2019 Nov;49:24. doi: 10.1016/j.ebiom.2019.09.056. Epub 2019 Oct 22. EBioMedicine. 2019. PMID: 31653586 Free PMC article. No abstract available.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
