

Comparative Genomics and Transcriptomics During Sexual Development Gives Insight Into the Life History of the Cosmopolitan Fungus Fusarium neocosmosporiellum
- PMID: 31231336
- PMCID: PMC6568001
- DOI: 10.3389/fmicb.2019.01247
Comparative Genomics and Transcriptomics During Sexual Development Gives Insight Into the Life History of the Cosmopolitan Fungus Fusarium neocosmosporiellum
Abstract
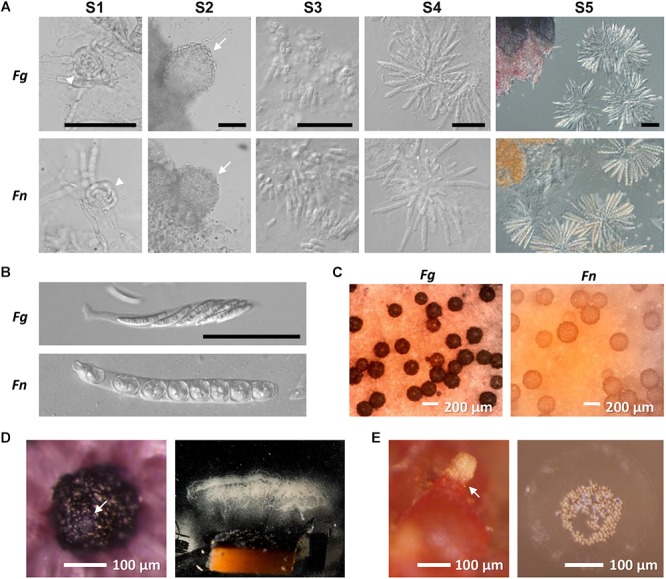
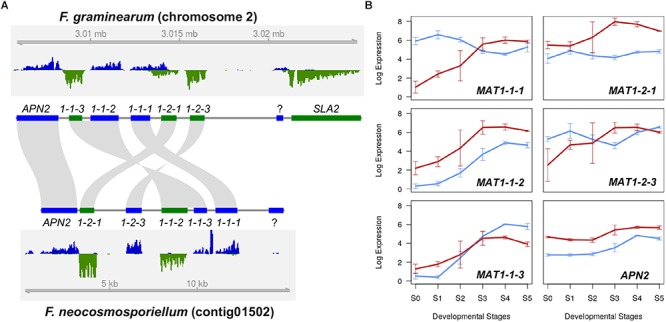
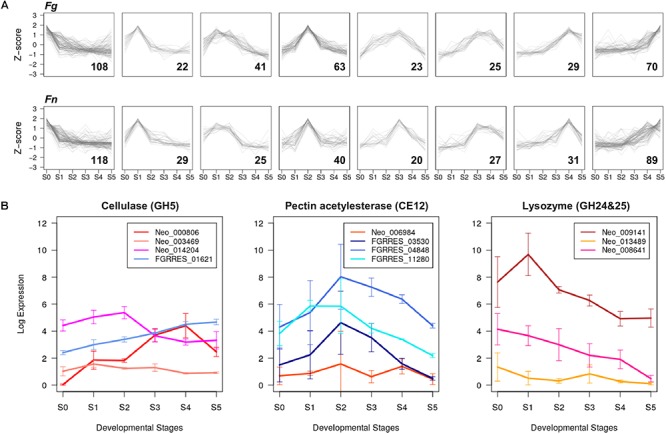
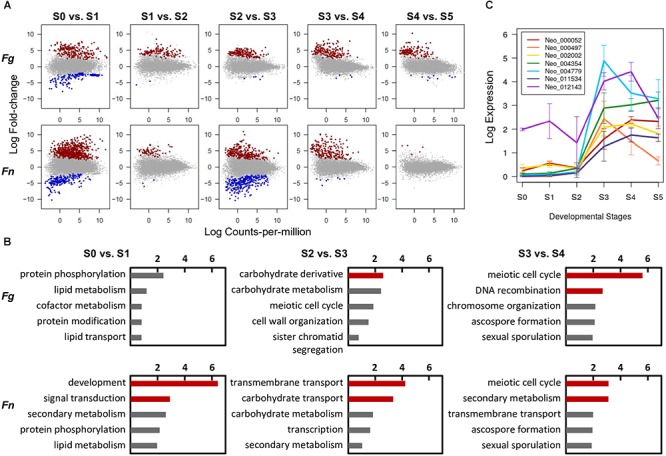
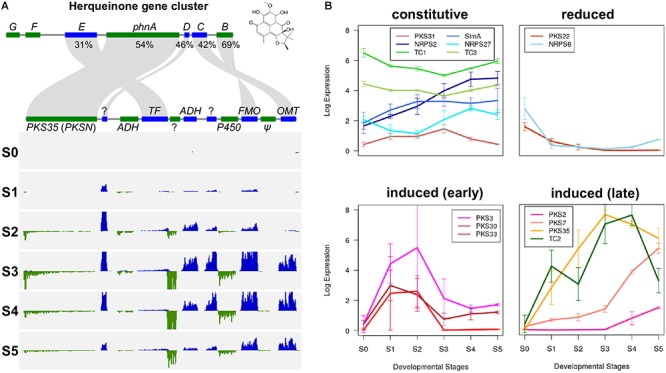
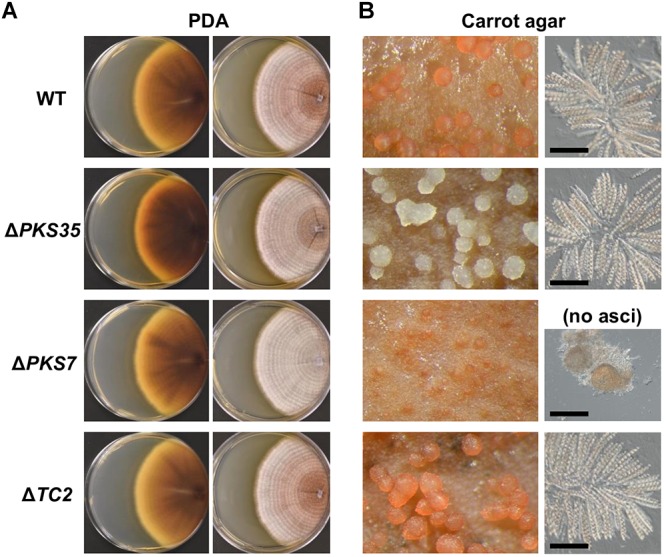
Fusarium neocosmosporiellum (formerly Neocosmospora vasinfecta) is a cosmopolitan fungus that has been reported from soil, herbivore dung, and as a fruit- and root-rot pathogen of numerous field crops, although it is not known to cause significant losses on any crop. Taking advantage of the fact that this species produces prolific numbers of perithecia in culture, the genome of F. neocosmosporiellum was sequenced and transcriptomic analysis across five stages of perithecium development was performed to better understand the metabolic potential for sexual development and gain insight into its life history. Perithecium morphology together with the genome and transcriptome were compared with those of the plant pathogen F. graminearum, a model for studying perithecium development. Larger ascospores of F. neocosmosporiellum and their tendency to discharge as a cluster demonstrated a duality of dispersal: the majority are passively dispersed through the formation of cirrhi, while a minority of spores are shot longer distances than those of F. graminearum. The predicted gene number in the F. neocosmosporiellum genome was similar to that in F. graminearum, but F. neocosmosporiellum had more carbohydrate metabolism-related and transmembrane transport genes. Many transporter genes were differentially expressed during perithecium development in F. neocosmosporiellum, which may account for its larger perithecia. Comparative analysis of the secondary metabolite gene clusters identified several polyketide synthase genes that were induced during later stages of perithecium development. Deletion of a polyketide synthase gene in F. neocosmosporiellum resulted in a defective perithecium phenotype, suggesting an important role of the corresponding metabolite, which has yet to be identified, in perithecium development. Results of this study have provided novel insights into the genomic underpinning of development in F. neocosmosporiellum, which may help elucidate its ability to occupy diverse ecological niches.
Keywords: Fusarium; mating type locus; perithecium; secondary metabolic genes; sexual development; transcriptome.
Figures
Similar articles
-
Sexual development and ascospore discharge in Fusarium graminearum.J Vis Exp. 2012 Mar 29;(61):3895. doi: 10.3791/3895. J Vis Exp. 2012. PMID: 22491175 Free PMC article.
-
The novel bZIP transcription factor Fpo1 negatively regulates perithecial development by modulating carbon metabolism in the ascomycete fungus Fusarium graminearum.Environ Microbiol. 2020 Jul;22(7):2596-2612. doi: 10.1111/1462-2920.14960. Epub 2020 Mar 5. Environ Microbiol. 2020. PMID: 32100421
-
FgPKS7 is an essential player in mating-type-mediated regulatory pathway required for completing sexual cycle in Fusarium graminearum.Environ Microbiol. 2021 Apr;23(4):1972-1990. doi: 10.1111/1462-2920.15305. Epub 2020 Nov 17. Environ Microbiol. 2021. PMID: 33169919
-
Recent advances in genes involved in secondary metabolite synthesis, hyphal development, energy metabolism and pathogenicity in Fusarium graminearum (teleomorph Gibberella zeae).Biotechnol Adv. 2014 Mar-Apr;32(2):390-402. doi: 10.1016/j.biotechadv.2013.12.007. Epub 2014 Jan 2. Biotechnol Adv. 2014. PMID: 24389085 Review.
-
Transcriptomics of cereal-Fusarium graminearum interactions: what we have learned so far.Mol Plant Pathol. 2018 Mar;19(3):764-778. doi: 10.1111/mpp.12561. Epub 2017 Jun 7. Mol Plant Pathol. 2018. PMID: 28411402 Free PMC article. Review.
Cited by
-
Whole-Genome Sequencing and Comparative Genome Analysis of Fusarium solani-melongenae Causing Fusarium Root and Stem Rot in Sweetpotatoes.Microbiol Spectr. 2022 Aug 31;10(4):e0068322. doi: 10.1128/spectrum.00683-22. Epub 2022 Jul 7. Microbiol Spectr. 2022. PMID: 35863027 Free PMC article.
-
Transcriptional Divergence Underpinning Sexual Development in the Fungal Class Sordariomycetes.mBio. 2022 Jun 28;13(3):e0110022. doi: 10.1128/mbio.01100-22. Epub 2022 May 31. mBio. 2022. PMID: 35638737 Free PMC article.
-
Transcription factor-dependent regulatory networks of sexual reproduction in Fusarium graminearum.mBio. 2025 Jan 8;16(1):e0303024. doi: 10.1128/mbio.03030-24. Epub 2024 Nov 26. mBio. 2025. PMID: 39589130 Free PMC article.
-
Fusarium: more than a node or a foot-shaped basal cell.Stud Mycol. 2021 Aug 17;98:100116. doi: 10.1016/j.simyco.2021.100116. eCollection 2021 Apr. Stud Mycol. 2021. PMID: 34466168 Free PMC article.
-
Fungal quinones: diversity, producers, and applications of quinones from Aspergillus, Penicillium, Talaromyces, Fusarium, and Arthrinium.Appl Microbiol Biotechnol. 2021 Nov;105(21-22):8157-8193. doi: 10.1007/s00253-021-11597-0. Epub 2021 Oct 9. Appl Microbiol Biotechnol. 2021. PMID: 34625822 Review.
References
-
- Aylor D. E., Anagnostakis S. L. (1991). Active discharge distance of ascospores of Venturia inaequalis. Phytopathology 81 548–551.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases