

Congenital lipodystrophy induces severe osteosclerosis
- PMID: 31233501
- PMCID: PMC6611650
- DOI: 10.1371/journal.pgen.1008244
Congenital lipodystrophy induces severe osteosclerosis
Abstract
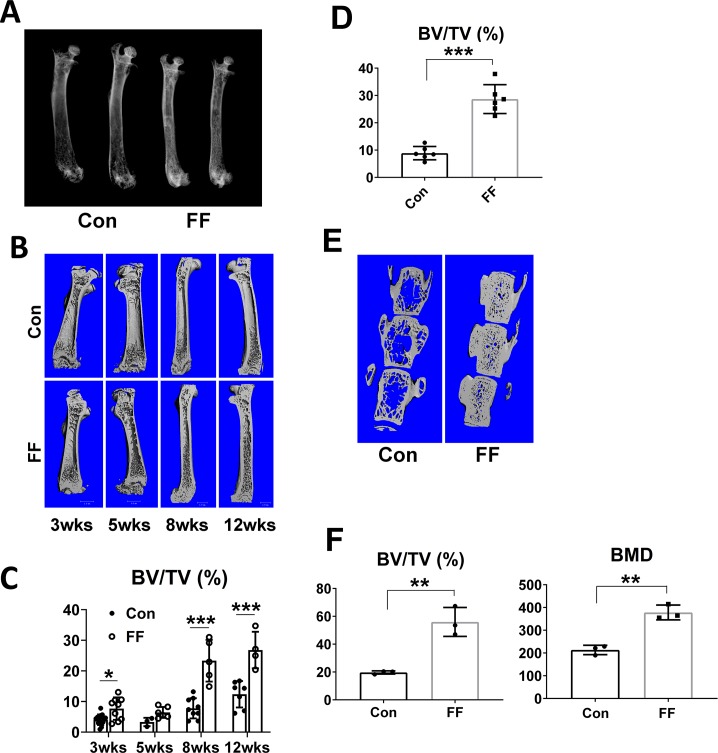
Berardinelli-Seip congenital generalized lipodystrophy is associated with increased bone mass suggesting that fat tissue regulates the skeleton. Because there is little mechanistic information regarding this issue, we generated "fat-free" (FF) mice completely lacking visible visceral, subcutaneous and brown fat. Due to robust osteoblastic activity, trabecular and cortical bone volume is markedly enhanced in these animals. FF mice, like Berardinelli-Seip patients, are diabetic but normalization of glucose tolerance and significant reduction in circulating insulin fails to alter their skeletal phenotype. Importantly, the skeletal phenotype of FF mice is completely rescued by transplantation of adipocyte precursors or white or brown fat depots, indicating that adipocyte derived products regulate bone mass. Confirming such is the case, transplantation of fat derived from adiponectin and leptin double knockout mice, unlike that obtained from their WT counterparts, fails to normalize FF bone. These observations suggest a paucity of leptin and adiponectin may contribute to the increased bone mass of Berardinelli-Seip patients.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
-
- Cohen A, Dempster DW, Recker RR, Lappe JM, Zhou H, Zwahlen A, et al. Abdominal fat is associated with lower bone formation and inferior bone quality in healthy premenopausal women: a transiliac bone biopsy study. J Clin Endocrinol Metab. 2013;98(6):2562–72. Epub 2013/03/22. 10.1210/jc.2013-1047 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
