

Stationary phase persister formation in Escherichia coli can be suppressed by piperacillin and PBP3 inhibition
- PMID: 31234796
- PMCID: PMC6591824
- DOI: 10.1186/s12866-019-1506-7
Stationary phase persister formation in Escherichia coli can be suppressed by piperacillin and PBP3 inhibition
Abstract
Background: Persisters are rare phenotypic variants within a bacterial population that are capable of tolerating lethal antibiotic concentrations. Passage through stationary phase is associated with the formation of persisters (type I), and a major physiological response of Escherichia coli during stationary phase is cell wall restructuring. Given the concurrence of these processes, we sought to assess whether perturbation to cell wall synthesis during stationary phase impacts type I persister formation.
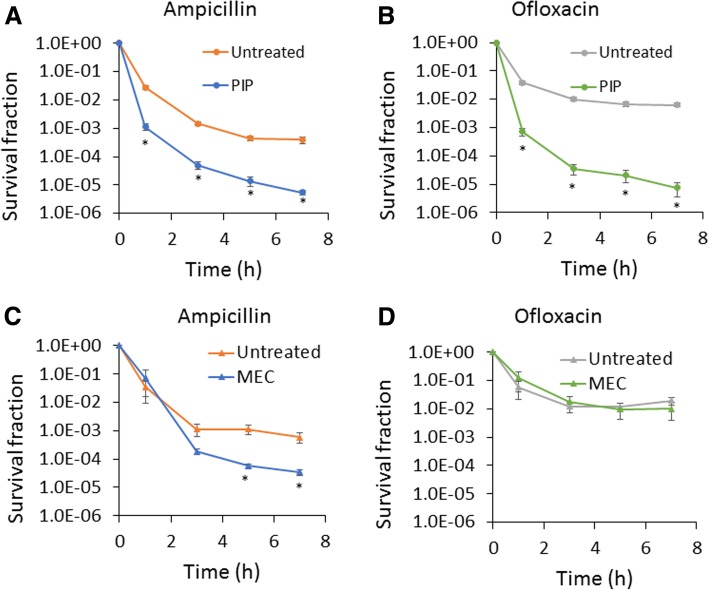
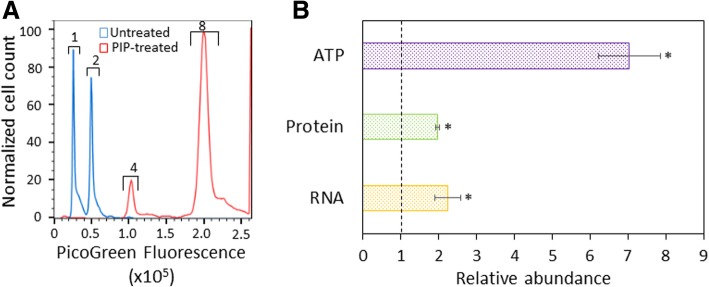
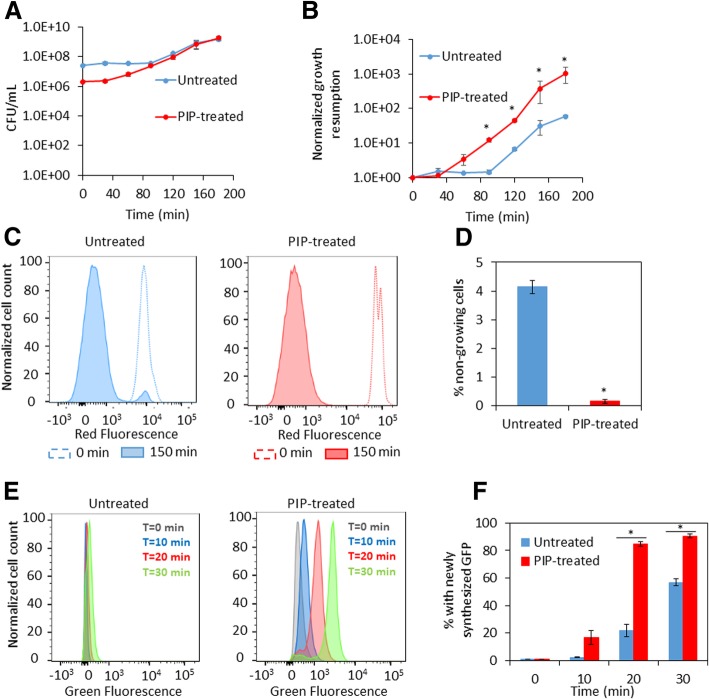
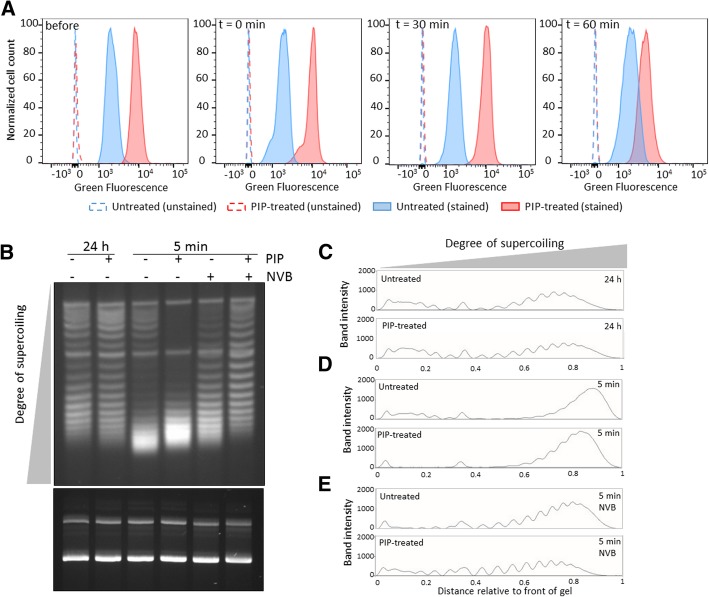
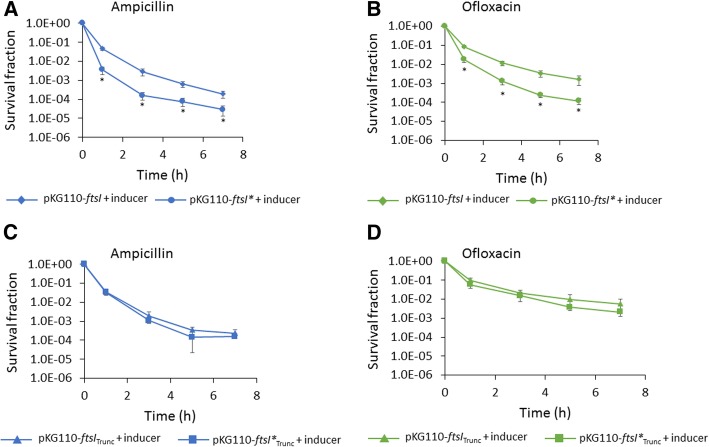
Results: We tested a panel of cell wall inhibitors and found that piperacillin, which primarily targets penicillin binding protein 3 (PBP3 encoded by ftsI), resulted in a significant reduction in both β-lactam (ampicillin, carbenicillin) and fluoroquinolone (ofloxacin, ciprofloxacin) persister levels. Further analyses showed that piperacillin exposure through stationary phase resulted in cells with more ATP, DNA, RNA, and protein (including PBPs) than untreated controls; and that their physiology led to more rapid resumption of DNA gyrase supercoiling activity, translation, and cell division upon introduction into fresh media. Previously, PBP3 inhibition had been linked to antibiotic efficacy through the DpiBA two component system; however, piperacillin suppressed persister formation in ΔdpiA to the same extent as it did in wild-type, suggesting that DpiBA is not required for the phenomenon reported here. To test the generality of PBP3 inhibition on persister formation, we expressed FtsI Ser307Ala to genetically inhibit PBP3, and suppression of persister formation was also observed, although not to the same magnitude as that seen for piperacillin treatment.
Conclusions: From these data we conclude that stationary phase PBP3 activity is important to type I persister formation in E. coli.
Keywords: E. coli; Fluoroquinolone; PBP3; Persisters; Piperacillin; β-Lactam.
Conflict of interest statement
None to declare.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
