

Master regulator analysis of paragangliomas carrying SDHx, VHL, or MAML3 genetic alterations
- PMID: 31234811
- PMCID: PMC6591808
- DOI: 10.1186/s12885-019-5813-z
Master regulator analysis of paragangliomas carrying SDHx, VHL, or MAML3 genetic alterations
Abstract
Background: Succinate dehydrogenase (SDH) loss and mastermind-like 3 (MAML3) translocation are two clinically important genetic alterations that correlate with increased rates of metastasis in subtypes of human paraganglioma and pheochromocytoma (PPGL) neuroendocrine tumors. Although hypotheses propose that succinate accumulation after SDH loss poisons dioxygenases and activates pseudohypoxia and epigenomic hypermethylation, it remains unclear whether these mechanisms account for oncogenic transcriptional patterns. Additionally, MAML3 translocation has recently been identified as a genetic alteration in PPGL, but is poorly understood. We hypothesize that a key to understanding tumorigenesis driven by these genetic alterations is identification of the transcription factors responsible for the observed oncogenic transcriptional changes.
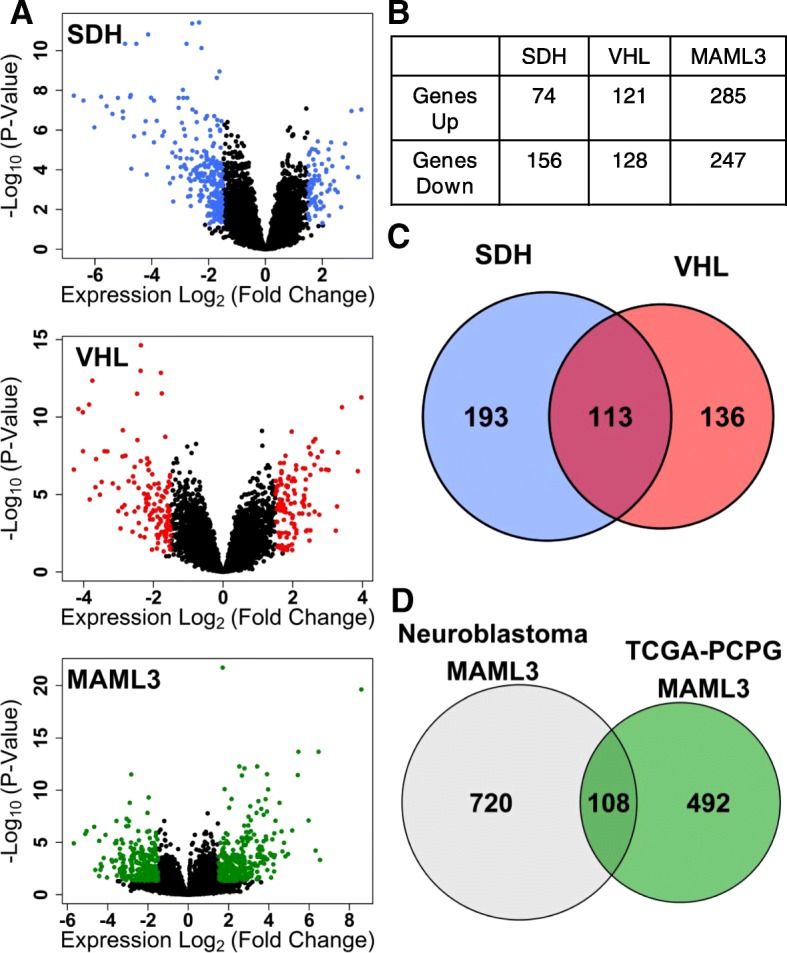
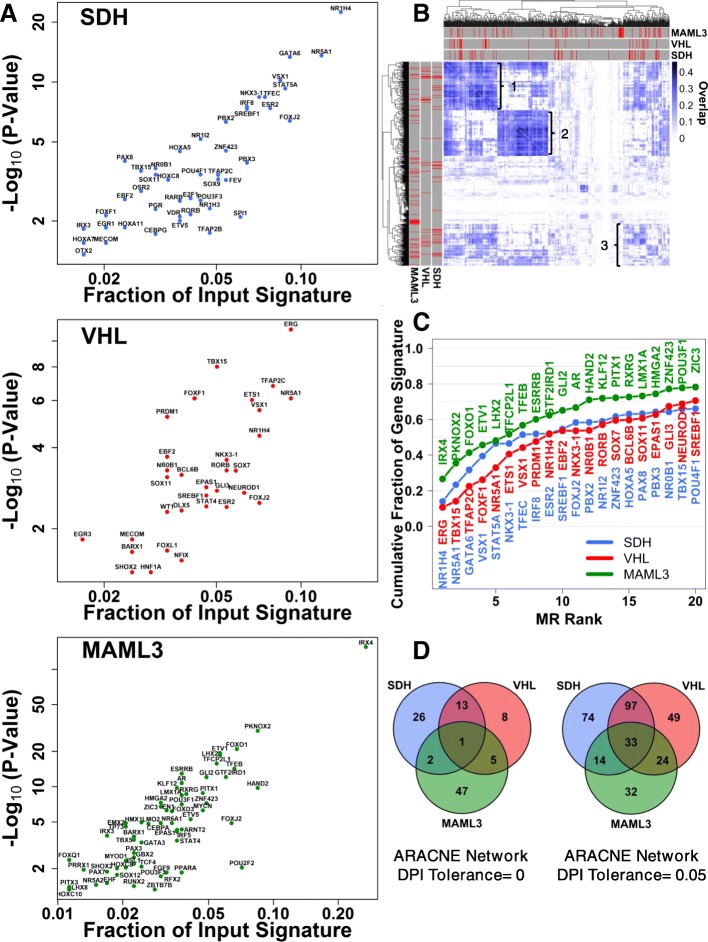
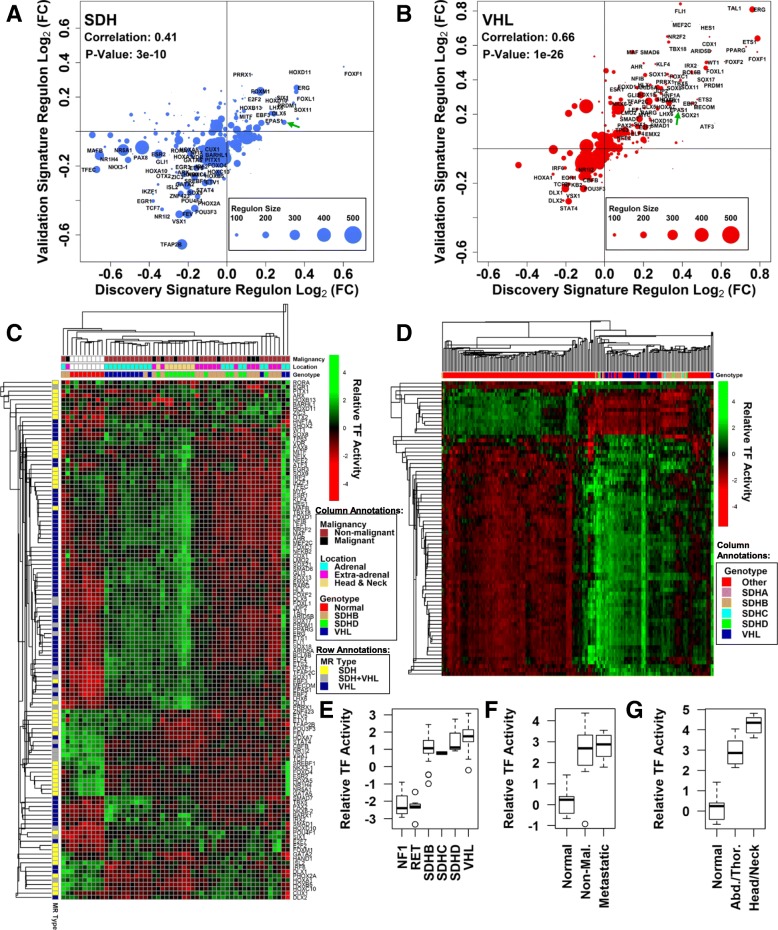
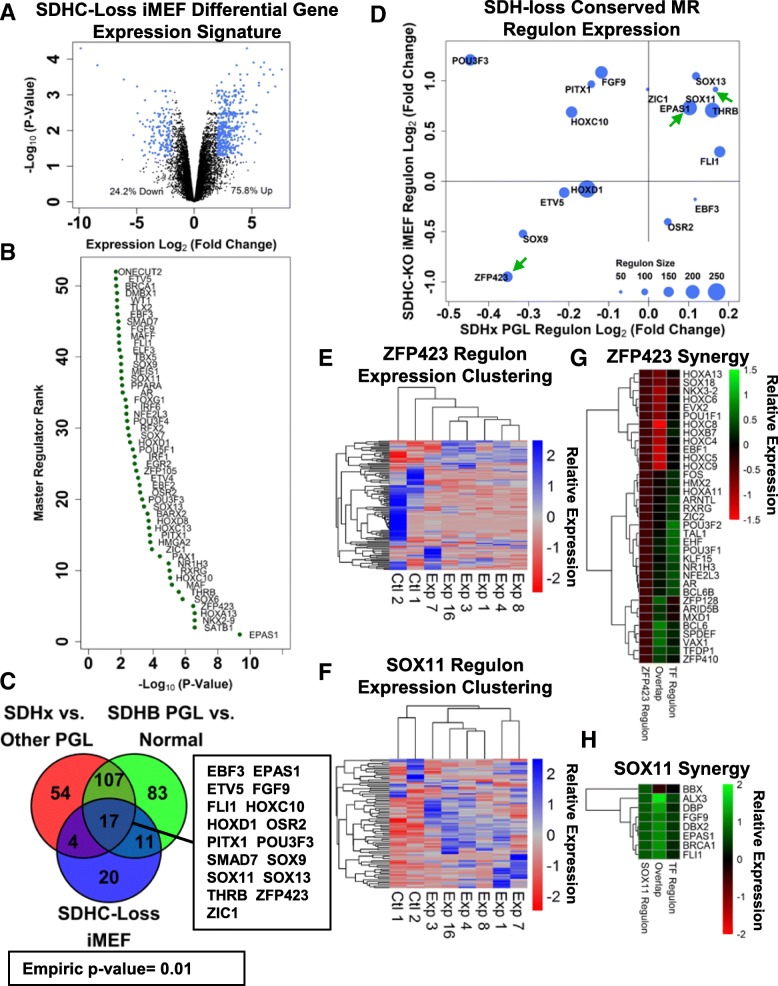
Methods: We leverage publicly-available human tumor gene expression profiling experiments (N = 179) to reconstruct a PPGL tumor-specific transcriptional network. We subsequently use the inferred transcriptional network to perform master regulator analyses nominating transcription factors predicted to control oncogenic transcription in specific PPGL molecular subtypes. Results are validated by analysis of an independent collection of PPGL tumor specimens (N = 188). We then perform a similar master regulator analysis in SDH-loss mouse embryonic fibroblasts (MEFs) to infer aspects of SDH loss master regulator response conserved across species and tissue types.
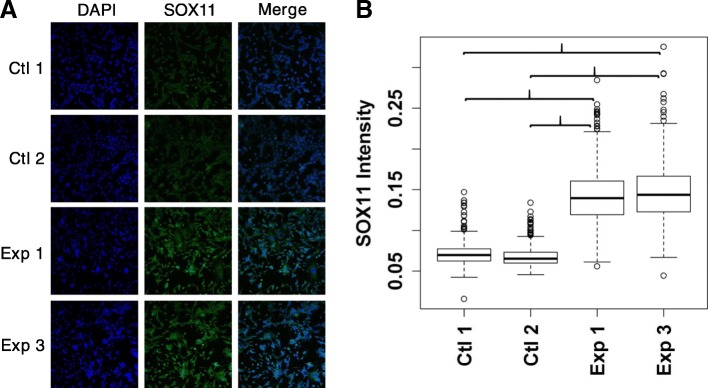
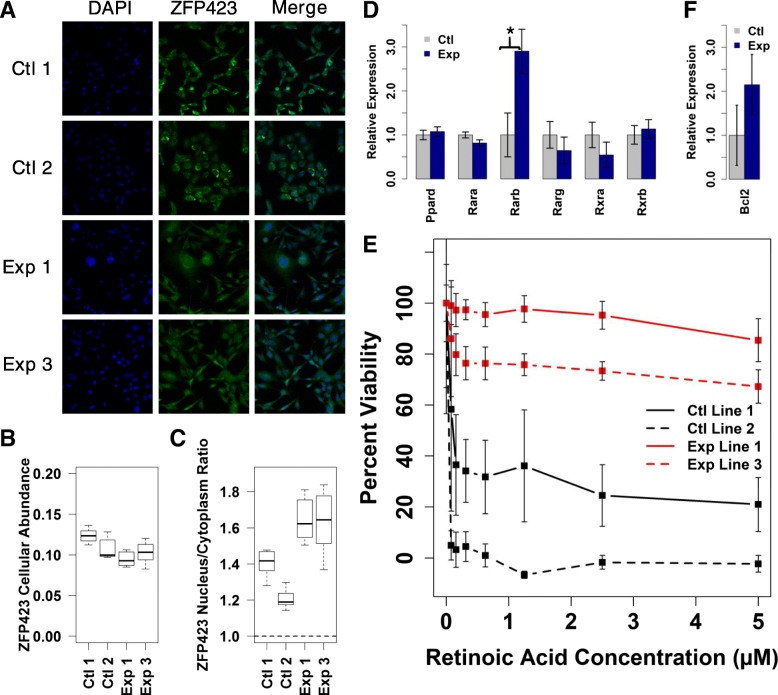
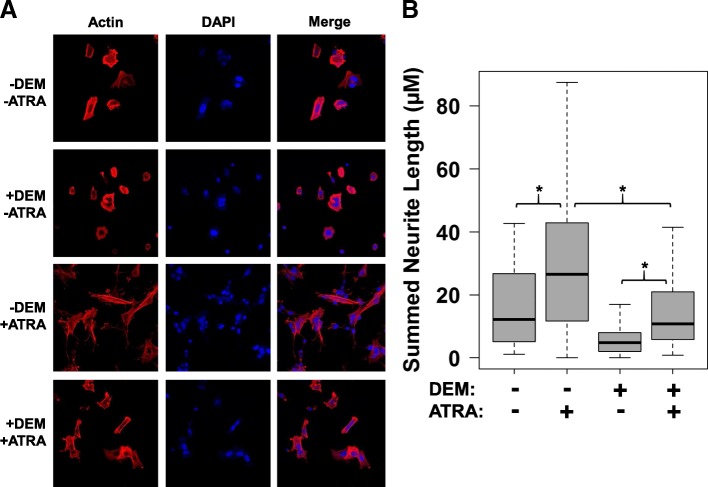
Results: A small number of master regulator transcription factors are predicted to drive the observed subtype-specific gene expression patterns in SDH loss and MAML3 translocation-positive PPGL. Interestingly, although EPAS1 perturbation is detectible in SDH-loss and VHL-loss tumors, it is by no means the most potent factor driving observed patterns of transcriptional dysregulation. Analysis of conserved SDH-loss master regulators in human tumors and MEFs implicated ZNF423, a known modulator of retinoic acid response in neuroblastoma. Subsequent functional analysis revealed a blunted cell death response to retinoic acid in SDH-loss MEFs and blunted differentiation response in SDH-inhibited SH-SY5Y neuroblastoma cells.
Conclusions: The unbiased analyses presented here nominate specific transcription factors that are likely drivers of oncogenic transcription in PPGL tumors. This information has the potential to be exploited for targeted therapy. Additionally, the observation that SDH loss or inhibition results in blunted retinoic acid response suggests a potential developmental etiology for this tumor subtype.
Keywords: Mastermind-like transcriptional coactivator 3; Paraganglioma; Pheochromocytoma; Retinoic acid; Succinate dehydrogenase; Transcription factor; Transcriptional network; von Hippel-Lindau.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Crona J, Delgado Verdugo A, Maharjan R, Stalberg P, Granberg D, Hellman P, Bjorklund P. Somatic mutations in H-RAS in sporadic pheochromocytoma and paraganglioma identified by exome sequencing. J Clin Endocrinol Metab. 2013;98(7):E1266–E1271. - PubMed
-
- Takahashi M, Ritz J, Cooper GM. Activation of a novel human transforming gene, ret, by DNA rearrangement. Cell. 1985;42(2):581–588. - PubMed
-
- Santoro M, Rosati R, Grieco M, Berlingieri MT, D'Amato GL, de Franciscis V, Fusco A. The ret proto-oncogene is consistently expressed in human pheochromocytomas and thyroid medullary carcinomas. Oncogene. 1990;5(10):1595–1598. - PubMed
-
- Wallace MR, Marchuk DA, Andersen LB, Letcher R, Odeh HM, Saulino AM, Fountain JW, Brereton A, Nicholson J, Mitchell AL, et al. Type 1 neurofibromatosis gene: identification of a large transcript disrupted in three NF1 patients. Science. 1990;249(4965):181–186. - PubMed
-
- Comino-Mendez I, Gracia-Aznarez FJ, Schiavi F, Landa I, Leandro-Garcia LJ, Leton R, Honrado E, Ramos-Medina R, Caronia D, Pita G, et al. Exome sequencing identifies MAX mutations as a cause of hereditary pheochromocytoma. Nat Genet. 2011;43(7):663–667. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
