

Functional Roles of Metabolic Intermediates in Regulating the Human Mitochondrial NAD(P)+-Dependent Malic Enzyme
- PMID: 31235710
- PMCID: PMC6591397
- DOI: 10.1038/s41598-019-45282-0
Functional Roles of Metabolic Intermediates in Regulating the Human Mitochondrial NAD(P)+-Dependent Malic Enzyme
Abstract
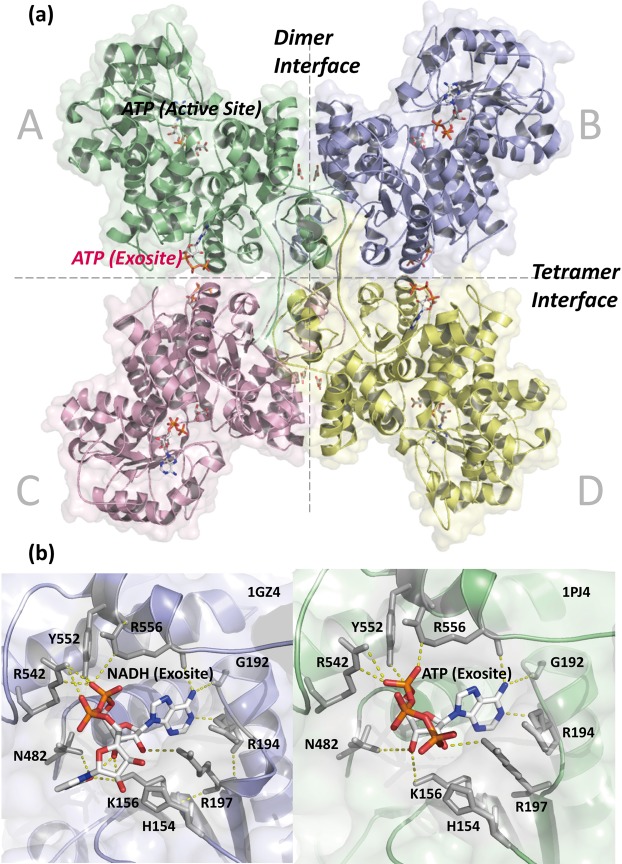
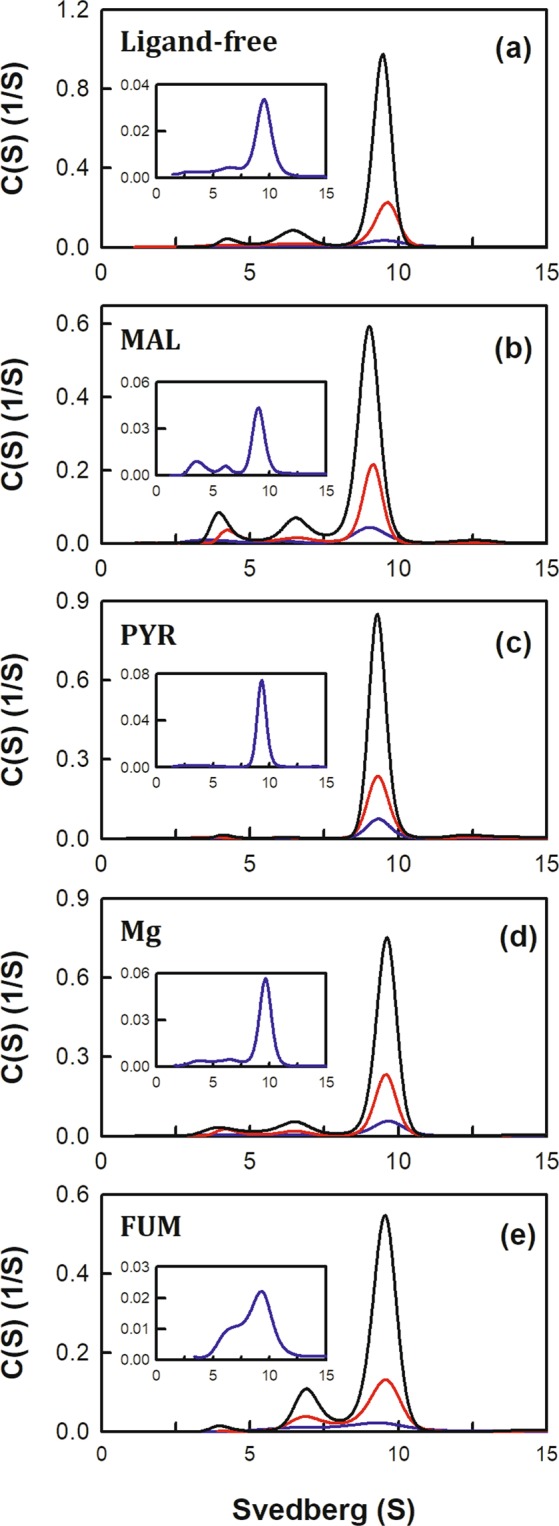
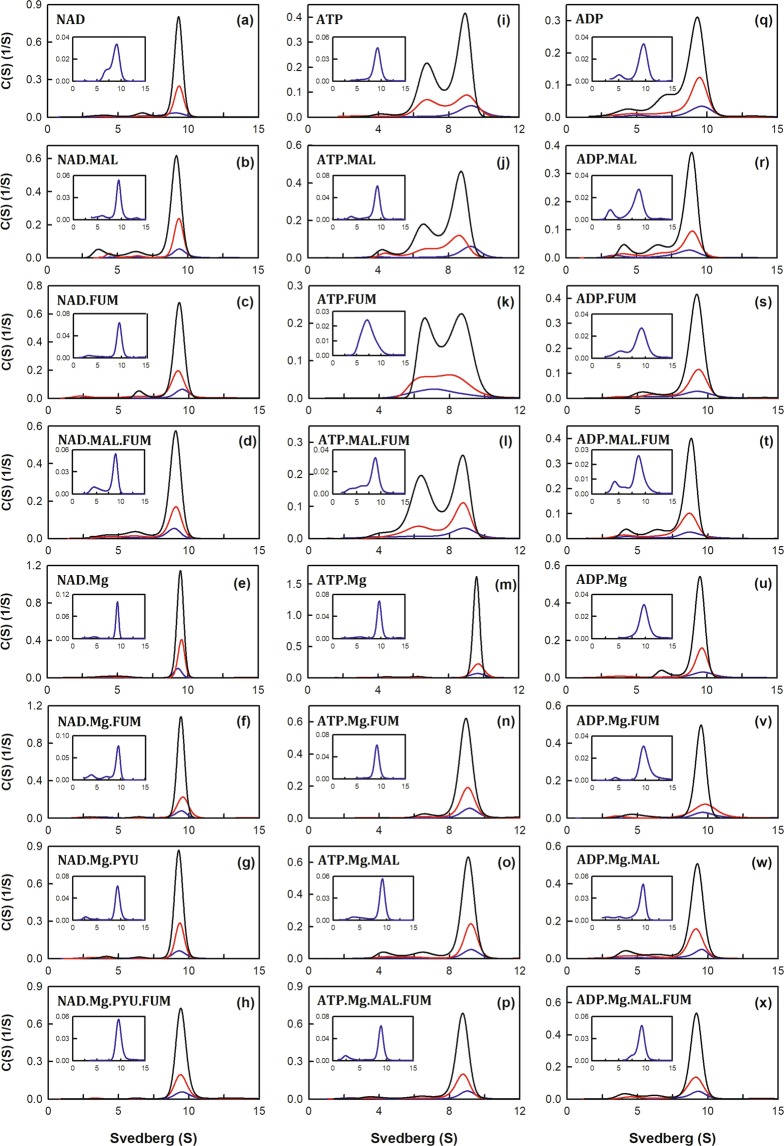
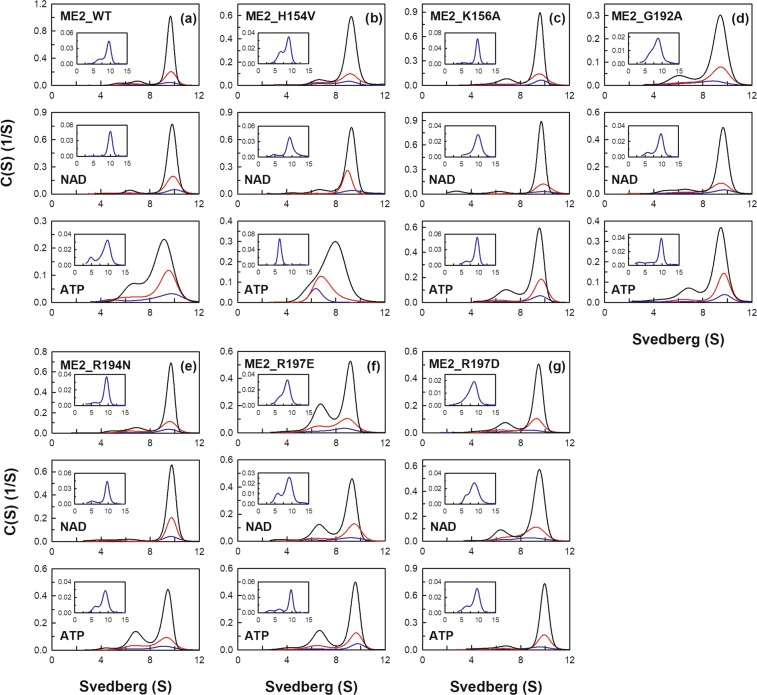
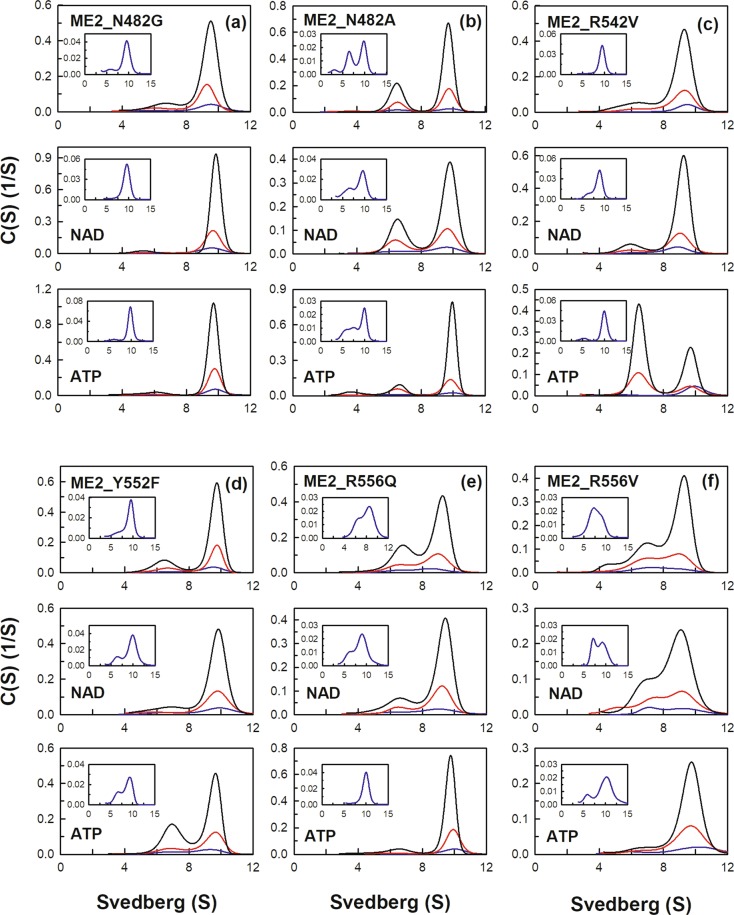
Human mitochondrial NAD(P)+-dependent malic enzyme (m-NAD(P)-ME) has a dimer of dimers quaternary structure with two independent allosteric sites in each monomer. Here, we reveal the different effects of nucleotide ligands on the quaternary structure regulation and functional role of the human m-NAD(P)-ME exosite. In this study, size distribution analysis was utilized to investigate the monomer-dimer-tetramer equilibrium of m-NAD(P)-ME in the presence of different ligands, and the monomer-dimer (Kd,12) and dimer-tetramer (Kd,24) dissociation constants were determined with these ligands. With NAD+, the enzyme formed more tetramers, and its Kd,24 (0.06 µM) was 6-fold lower than the apoenzyme Kd,24 (0.34 µM). When ATP was present, the enzyme displayed more dimers, and its Kd,24 (2.74 µM) was 8-fold higher than the apoenzyme. Similar to the apoenzyme, the ADP-bound enzyme was present as a tetramer with a small amount of dimers and monomers. These results indicate that NAD+ promotes association of the dimeric enzyme into tetramers, whereas ATP stimulates dissociation of the tetrameric enzyme into dimers, and ADP has little effect on the tetrameric stability of the enzyme. A series of exosite mutants were created using site-directed mutagenesis. Size distribution analysis and kinetic studies of these mutants with NAD+ or ATP indicated that Arg197, Asn482 and Arg556 are essential for the ATP binding and ATP-induced dissociation of human m-NAD(P)-ME. In summary, the present results demonstrate that nucleotides perform discrete functions regulating the quaternary structure and catalysis of m-NAD(P)-ME. Such regulation by the binding of different nucleotides may be critically associated with the physiological concentrations of these ligands.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Functional roles of the tetramer organization of malic enzyme.J Biol Chem. 2009 Jul 3;284(27):18096-105. doi: 10.1074/jbc.M109.005082. Epub 2009 May 5. J Biol Chem. 2009. PMID: 19416979 Free PMC article.
-
Dual roles of Lys(57) at the dimer interface of human mitochondrial NAD(P)+-dependent malic enzyme.Biochem J. 2009 May 13;420(2):201-9. doi: 10.1042/BJ20090076. Biochem J. 2009. PMID: 19236308
-
Dual functional roles of ATP in the human mitochondrial malic enzyme.Biochemistry. 2004 Jun 15;43(23):7382-90. doi: 10.1021/bi049600r. Biochemistry. 2004. PMID: 15182181
-
Malate decarboxylases: evolution and roles of NAD(P)-ME isoforms in species performing C(4) and C(3) photosynthesis.J Exp Bot. 2011 May;62(9):3061-9. doi: 10.1093/jxb/err024. Epub 2011 Mar 31. J Exp Bot. 2011. PMID: 21459769 Review.
-
Malate dehydrogenases--structure and function.Gen Physiol Biophys. 2002 Sep;21(3):257-65. Gen Physiol Biophys. 2002. PMID: 12537350 Review.
Cited by
-
Suppression of the human malic enzyme 2 modifies energy metabolism and inhibits cellular respiration.Commun Biol. 2023 May 22;6(1):548. doi: 10.1038/s42003-023-04930-y. Commun Biol. 2023. PMID: 37217557 Free PMC article.
-
Redox regulation of the immune response.Cell Mol Immunol. 2022 Oct;19(10):1079-1101. doi: 10.1038/s41423-022-00902-0. Epub 2022 Sep 2. Cell Mol Immunol. 2022. PMID: 36056148 Free PMC article. Review.
-
Lipid Peroxidation-Related Redox Signaling in Osteosarcoma.Int J Mol Sci. 2024 Apr 22;25(8):4559. doi: 10.3390/ijms25084559. Int J Mol Sci. 2024. PMID: 38674143 Free PMC article. Review.
-
Structural and Molecular Dynamics of Mycobacterium tuberculosis Malic Enzyme, a Potential Anti-TB Drug Target.ACS Infect Dis. 2021 Jan 8;7(1):174-188. doi: 10.1021/acsinfecdis.0c00735. Epub 2020 Dec 23. ACS Infect Dis. 2021. PMID: 33356117 Free PMC article.
-
Integrative analysis of toxicometabolomics and toxicoproteomics data: new molecular insights into thiazolidinedione-induced cardiotoxicity.Metabolomics. 2024 Dec 4;21(1):1. doi: 10.1007/s11306-024-02201-3. Metabolomics. 2024. PMID: 39636558 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
