

Structure of F1-ATPase from the obligate anaerobe Fusobacterium nucleatum
- PMID: 31238823
- PMCID: PMC6597759
- DOI: 10.1098/rsob.190066
Structure of F1-ATPase from the obligate anaerobe Fusobacterium nucleatum
Abstract
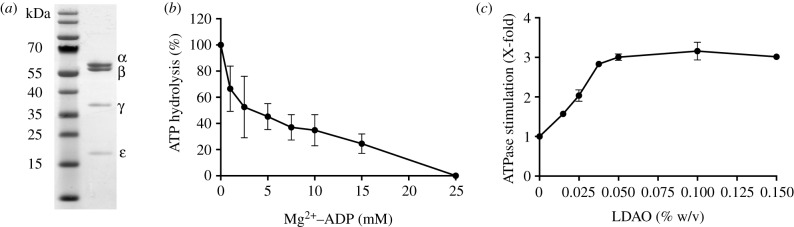
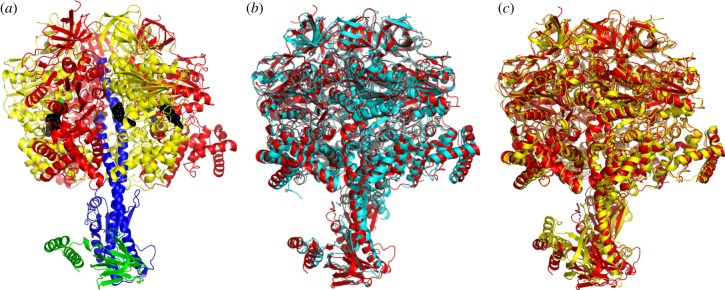
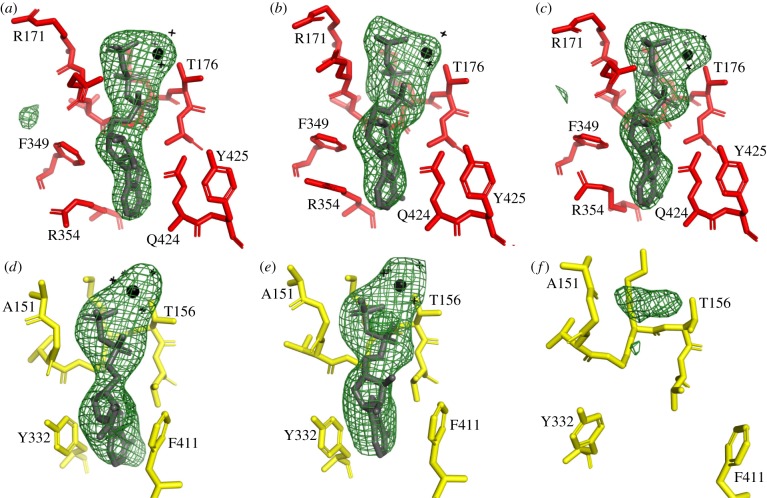
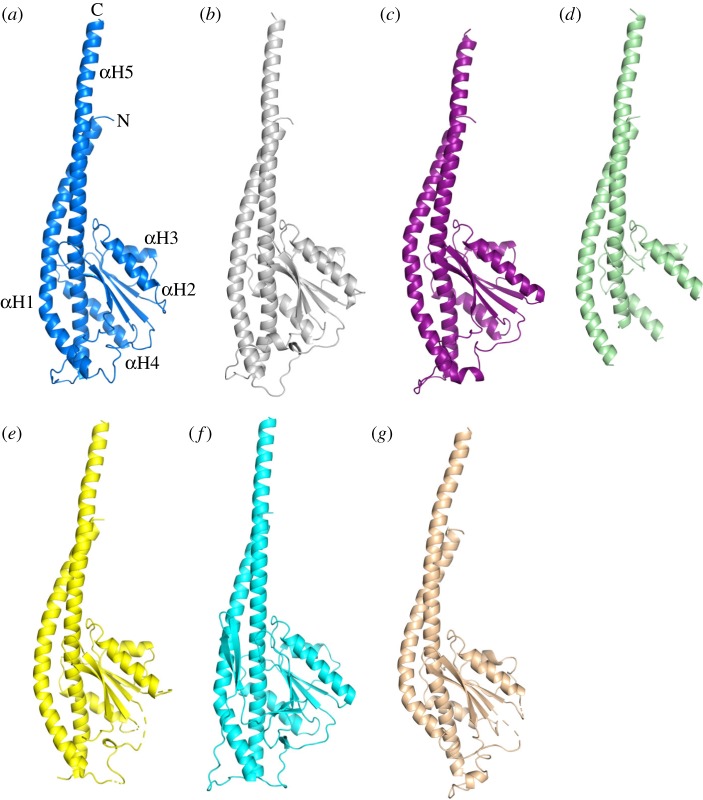
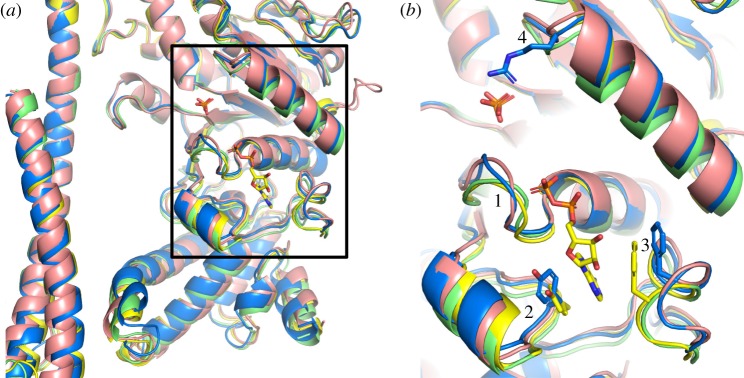
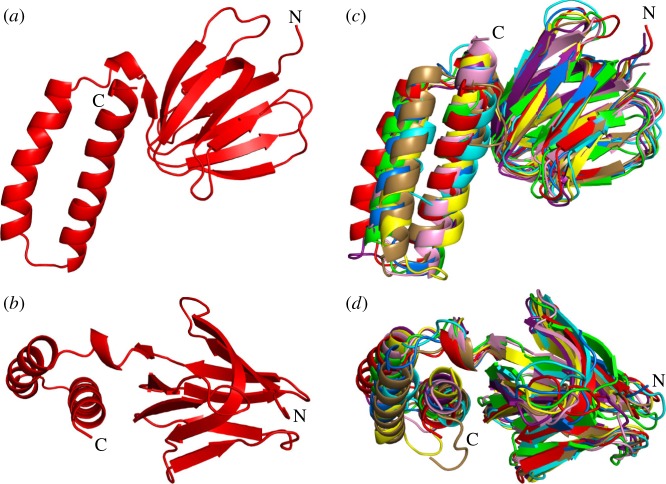
The crystal structure of the F1-catalytic domain of the adenosine triphosphate (ATP) synthase has been determined from the pathogenic anaerobic bacterium Fusobacterium nucleatum. The enzyme can hydrolyse ATP but is partially inhibited. The structure is similar to those of the F1-ATPases from Caldalkalibacillus thermarum, which is more strongly inhibited in ATP hydrolysis, and in Mycobacterium smegmatis, which has a very low ATP hydrolytic activity. The βE-subunits in all three enzymes are in the conventional 'open' state, and in the case of C. thermarum and M. smegmatis, they are occupied by an ADP and phosphate (or sulfate), but in F. nucleatum, the occupancy by ADP appears to be partial. It is likely that the hydrolytic activity of the F. nucleatum enzyme is regulated by the concentration of ADP, as in mitochondria.
Keywords: ATP hydrolysis; Fusobacterium nucleatum; catalytic F1-ATPase; pathogen; regulation; structure.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Regulation of the thermoalkaliphilic F1-ATPase from Caldalkalibacillus thermarum.Proc Natl Acad Sci U S A. 2016 Sep 27;113(39):10860-5. doi: 10.1073/pnas.1612035113. Epub 2016 Sep 12. Proc Natl Acad Sci U S A. 2016. PMID: 27621435 Free PMC article.
-
ADP-inhibited structure of non-catalytic site-depleted FoF1-ATPase from thermophilic Bacillus sp. PS-3.Biochim Biophys Acta Bioenerg. 2025 Apr 1;1866(2):149536. doi: 10.1016/j.bbabio.2025.149536. Epub 2025 Jan 7. Biochim Biophys Acta Bioenerg. 2025. PMID: 39788275
-
The structure of the catalytic domain of the ATP synthase from Mycobacterium smegmatis is a target for developing antitubercular drugs.Proc Natl Acad Sci U S A. 2019 Mar 5;116(10):4206-4211. doi: 10.1073/pnas.1817615116. Epub 2019 Jan 25. Proc Natl Acad Sci U S A. 2019. PMID: 30683723 Free PMC article.
-
Does the gamma subunit move to an abortive position of ATP hydrolysis when the F1.ADP.Mg complex isomerizes to the inactive F1*.ADP.Mg complex?J Bioenerg Biomembr. 1996 Oct;28(5):433-8. doi: 10.1007/BF02113985. J Bioenerg Biomembr. 1996. PMID: 8951090 Review.
-
Insights into the regulatory function of the ɛ subunit from bacterial F-type ATP synthases: a comparison of structural, biochemical and biophysical data.Open Biol. 2018 May;8(5):170275. doi: 10.1098/rsob.170275. Open Biol. 2018. PMID: 29769322 Free PMC article. Review.
Cited by
-
Hindguts of Kyphosus sydneyanus harbor phylogenetically and genomically distinct Alistipes capable of degrading algal polysaccharides and diazotrophy.mSystems. 2025 Jan 21;10(1):e0100724. doi: 10.1128/msystems.01007-24. Epub 2024 Dec 23. mSystems. 2025. PMID: 39714211 Free PMC article.
-
F1·Fo ATP Synthase/ATPase: Contemporary View on Unidirectional Catalysis.Int J Mol Sci. 2023 Mar 12;24(6):5417. doi: 10.3390/ijms24065417. Int J Mol Sci. 2023. PMID: 36982498 Free PMC article. Review.
References
-
- Walker JE. 2017. Structure, mechanism and regulation of ATP synthases. In Mechanisms of primary energy transduction in biology (ed. Wikström M.), pp. 338–373. Cambridge, UK: Royal Society of Chemistry.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
