

Identification of Extracellular DNA-Binding Proteins in the Biofilm Matrix
- PMID: 31239382
- PMCID: PMC6593408
- DOI: 10.1128/mBio.01137-19
Identification of Extracellular DNA-Binding Proteins in the Biofilm Matrix
Abstract
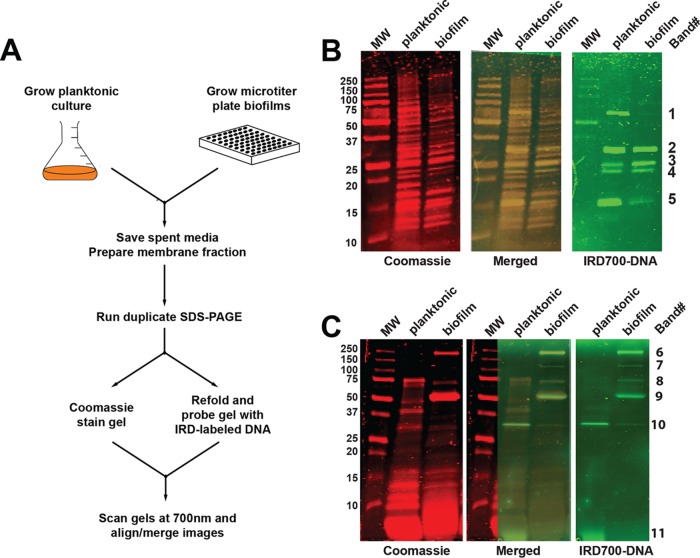
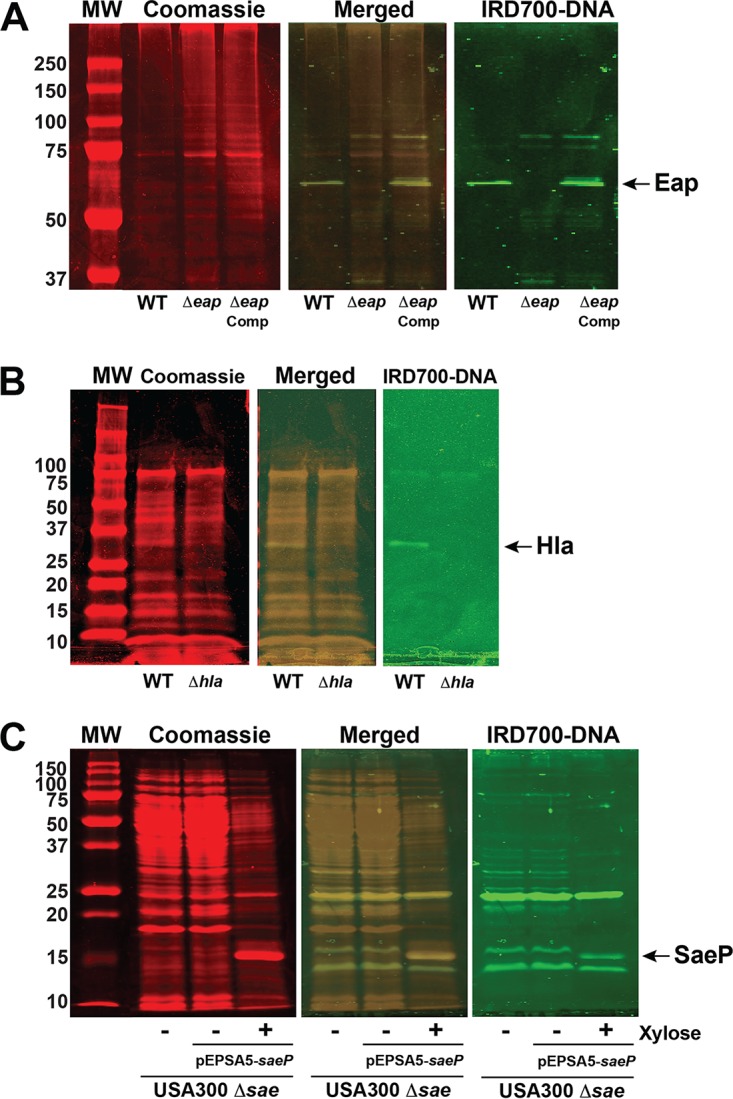
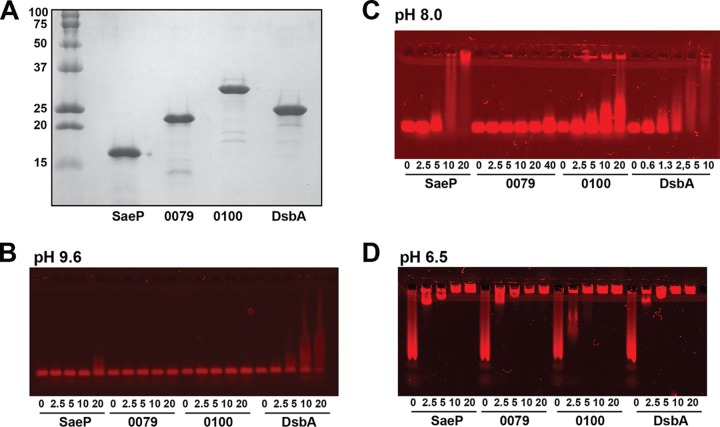
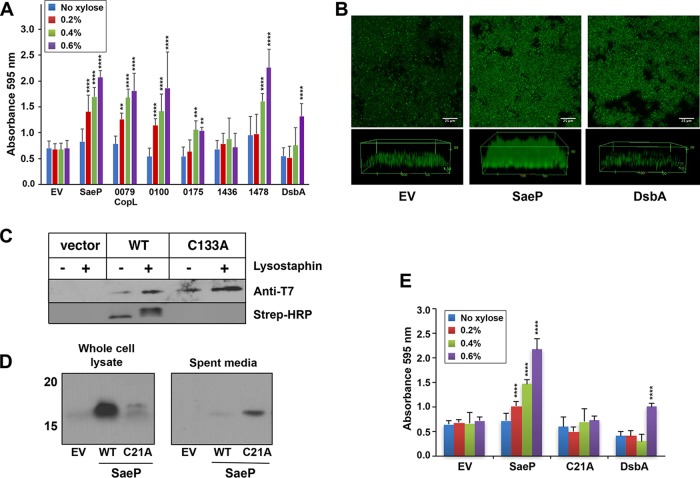
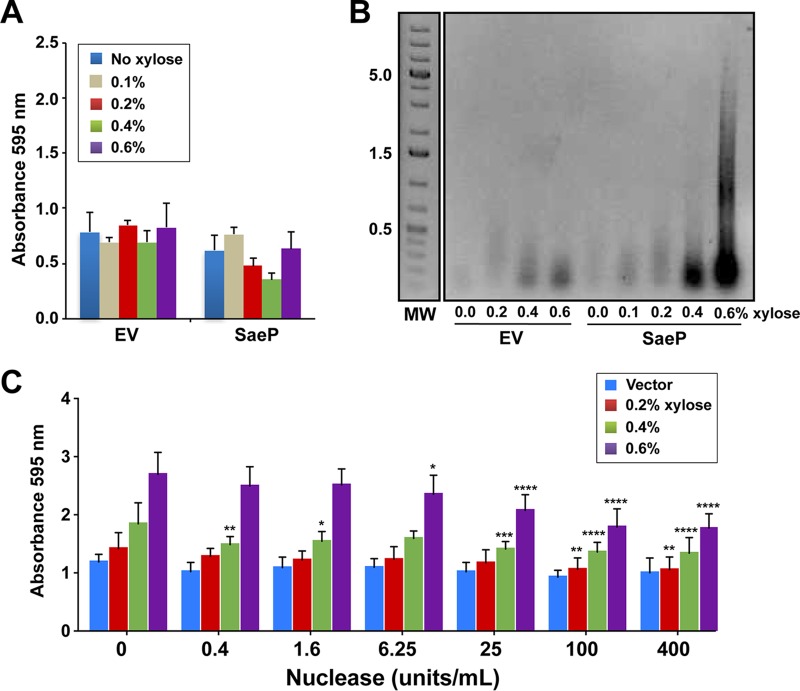
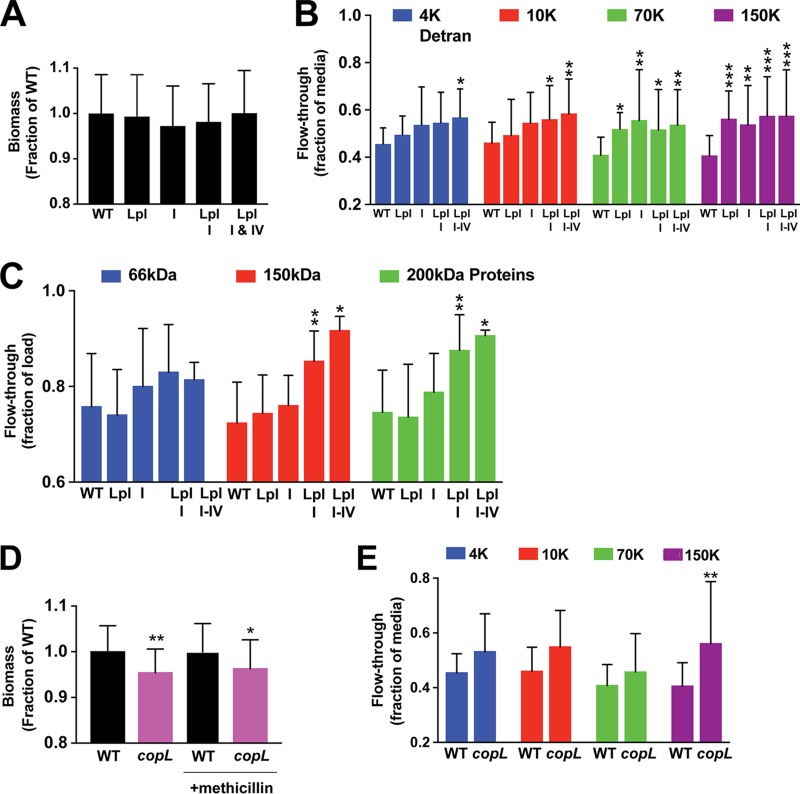
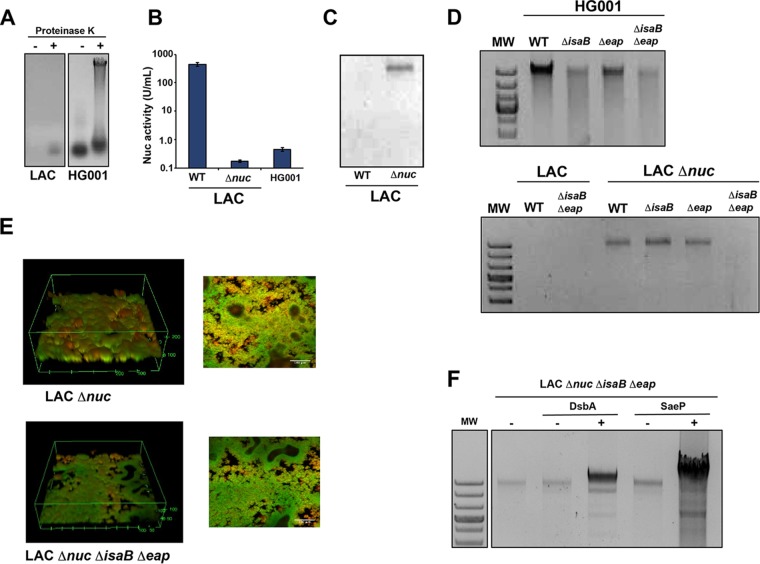
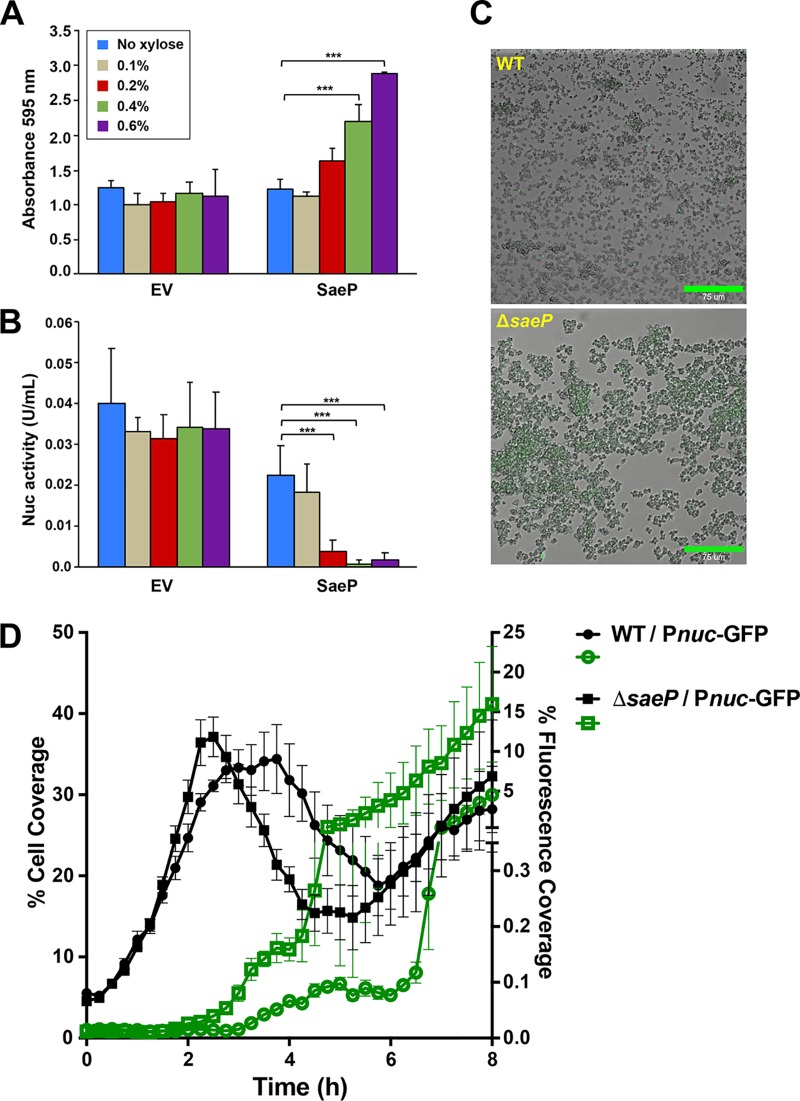
We developed a new approach that couples Southwestern blotting and mass spectrometry to discover proteins that bind extracellular DNA (eDNA) in bacterial biofilms. Using Staphylococcus aureus as a model pathogen, we identified proteins with known DNA-binding activity and uncovered a series of lipoproteins with previously unrecognized DNA-binding activity. We demonstrated that expression of these lipoproteins results in an eDNA-dependent biofilm enhancement. Additionally, we found that while deletion of lipoproteins had a minimal impact on biofilm accumulation, these lipoprotein mutations increased biofilm porosity, suggesting that lipoproteins and their associated interactions contribute to biofilm structure. For one of the lipoproteins, SaeP, we showed that the biofilm phenotype requires the lipoprotein to be anchored to the outside of the cellular membrane, and we further showed that increased SaeP expression correlates with more retention of high-molecular-weight DNA on the bacterial cell surface. SaeP is a known auxiliary protein of the SaeRS system, and we also demonstrated that the levels of SaeP correlate with nuclease production, which can further impact biofilm development. It has been reported that S. aureus biofilms are stabilized by positively charged cytoplasmic proteins that are released into the extracellular environment, where they make favorable electrostatic interactions with the negatively charged cell surface and eDNA. In this work we extend this electrostatic net model to include secreted eDNA-binding proteins and membrane-attached lipoproteins that can function as anchor points between eDNA in the biofilm matrix and the bacterial cell surface.IMPORTANCE Many bacteria are capable of forming biofilms encased in a matrix of self-produced extracellular polymeric substances (EPS) that protects them from chemotherapies and the host defenses. As a result of these inherent resistance mechanisms, bacterial biofilms are extremely difficult to eradicate and are associated with chronic wounds, orthopedic and surgical wound infections, and invasive infections, such as infective endocarditis and osteomyelitis. It is therefore important to understand the nature of the interactions between the bacterial cell surface and EPS that stabilize biofilms. Extracellular DNA (eDNA) has been recognized as an EPS constituent for many bacterial species and has been shown to be important in promoting biofilm formation. Using Staphylococcus aureus biofilms, we show that membrane-attached lipoproteins can interact with the eDNA in the biofilm matrix and promote biofilm formation, which suggests that lipoproteins are potential targets for novel therapies aimed at disrupting bacterial biofilms.
Keywords: MRSA; Southwestern blotting; Staphylococcus aureus; biofilms; eDNA; extracellular DNA; nuclease.
Copyright © 2019 Kavanaugh et al.
Figures
Similar articles
-
Genetic and Biochemical Analysis of CodY-Mediated Cell Aggregation in Staphylococcus aureus Reveals an Interaction between Extracellular DNA and Polysaccharide in the Extracellular Matrix.J Bacteriol. 2020 Mar 26;202(8):e00593-19. doi: 10.1128/JB.00593-19. Print 2020 Mar 26. J Bacteriol. 2020. PMID: 32015143 Free PMC article.
-
An Electrostatic Net Model for the Role of Extracellular DNA in Biofilm Formation by Staphylococcus aureus.J Bacteriol. 2015 Dec;197(24):3779-87. doi: 10.1128/JB.00726-15. Epub 2015 Sep 28. J Bacteriol. 2015. PMID: 26416831 Free PMC article.
-
Beyond the double helix: the multifaceted landscape of extracellular DNA in Staphylococcus aureus biofilms.Front Cell Infect Microbiol. 2024 Jun 5;14:1400648. doi: 10.3389/fcimb.2024.1400648. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38903938 Free PMC article. Review.
-
Glycosyltransferase-Mediated Biofilm Matrix Dynamics and Virulence of Streptococcus mutans.Appl Environ Microbiol. 2019 Feb 20;85(5):e02247-18. doi: 10.1128/AEM.02247-18. Print 2019 Mar 1. Appl Environ Microbiol. 2019. PMID: 30578260 Free PMC article.
-
Extracellular DNA (eDNA). A Major Ubiquitous Element of the Bacterial Biofilm Architecture.Int J Mol Sci. 2021 Aug 23;22(16):9100. doi: 10.3390/ijms22169100. Int J Mol Sci. 2021. PMID: 34445806 Free PMC article. Review.
Cited by
-
Biofilms in Periprosthetic Orthopedic Infections Seen through the Eyes of Neutrophils: How Can We Help Neutrophils?Int J Mol Sci. 2023 Nov 23;24(23):16669. doi: 10.3390/ijms242316669. Int J Mol Sci. 2023. PMID: 38068991 Free PMC article. Review.
-
Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies.Microbiol Mol Biol Rev. 2020 Aug 12;84(3):e00026-19. doi: 10.1128/MMBR.00026-19. Print 2020 Aug 19. Microbiol Mol Biol Rev. 2020. PMID: 32792334 Free PMC article. Review.
-
Systematic Full-Cycle Engineering Microbial Biofilms to Boost Electricity Production in Shewanella oneidensis.Research (Wash D C). 2023;6:0081. doi: 10.34133/research.0081. Epub 2023 Mar 15. Research (Wash D C). 2023. PMID: 36939407 Free PMC article.
-
Extracellular DNA: A Critical Aspect of Marine Biofilms.Microorganisms. 2022 Jun 24;10(7):1285. doi: 10.3390/microorganisms10071285. Microorganisms. 2022. PMID: 35889003 Free PMC article.
-
Different Polymers for the Base of Removable Dentures? Part II: A Narrative Review of the Dynamics of Microbial Plaque Formation on Dentures.Polymers (Basel). 2023 Dec 21;16(1):40. doi: 10.3390/polym16010040. Polymers (Basel). 2023. PMID: 38201705 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
