

Lzts1 controls both neuronal delamination and outer radial glial-like cell generation during mammalian cerebral development
- PMID: 31239441
- PMCID: PMC6592889
- DOI: 10.1038/s41467-019-10730-y
Lzts1 controls both neuronal delamination and outer radial glial-like cell generation during mammalian cerebral development
Abstract
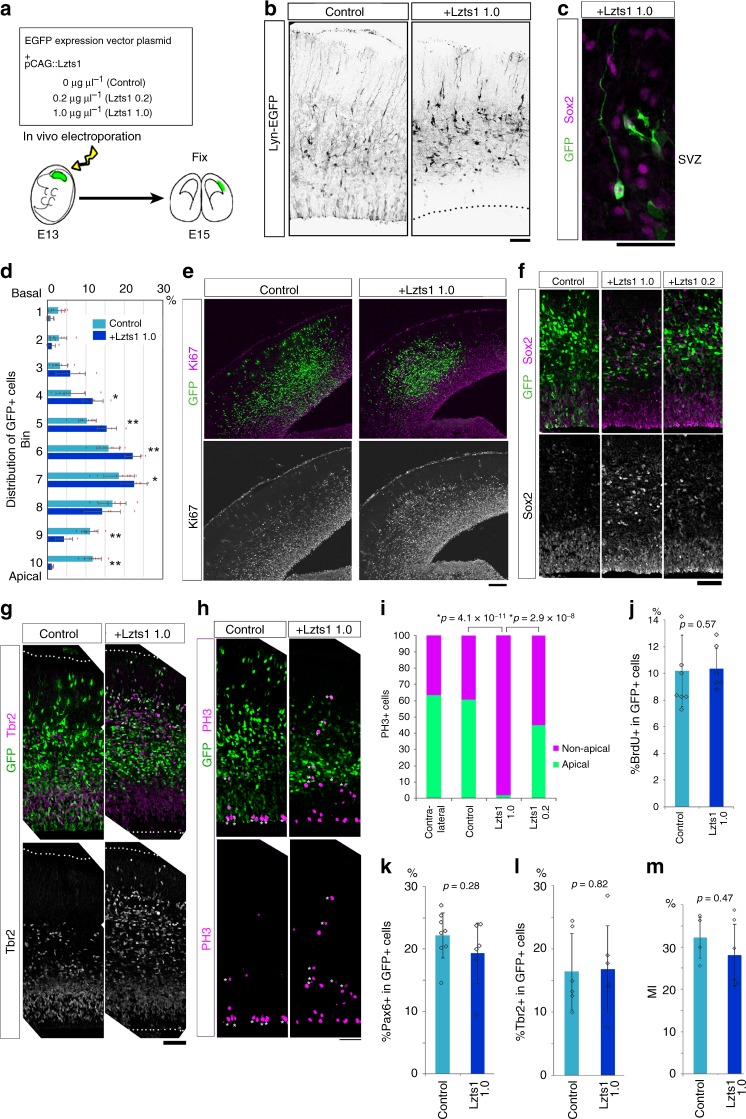
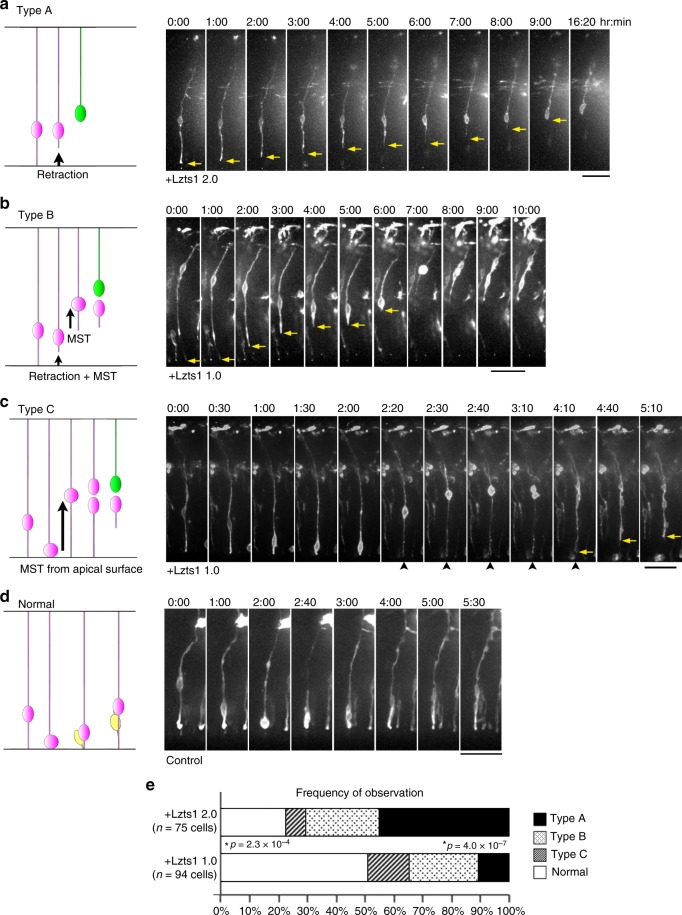
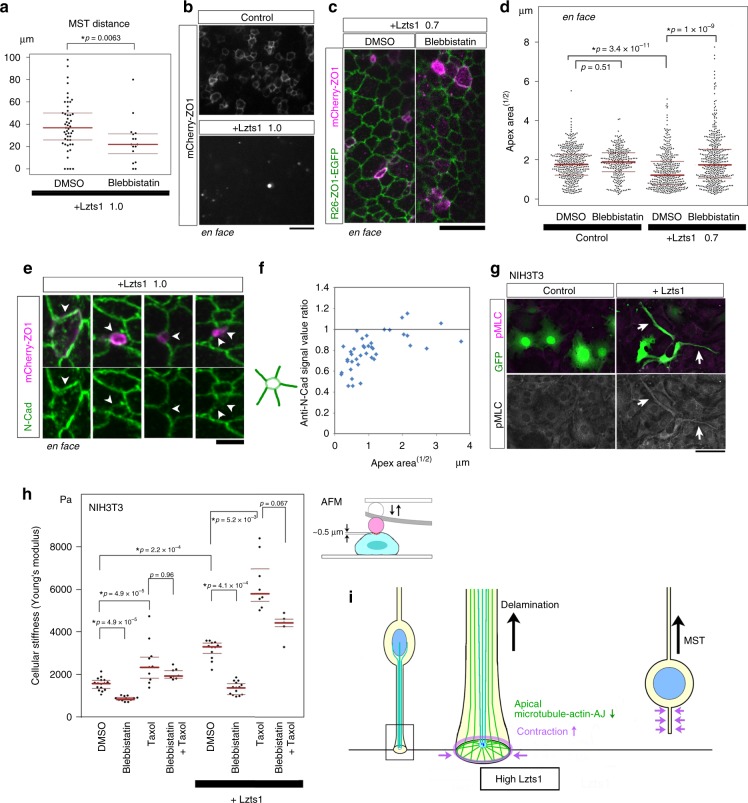
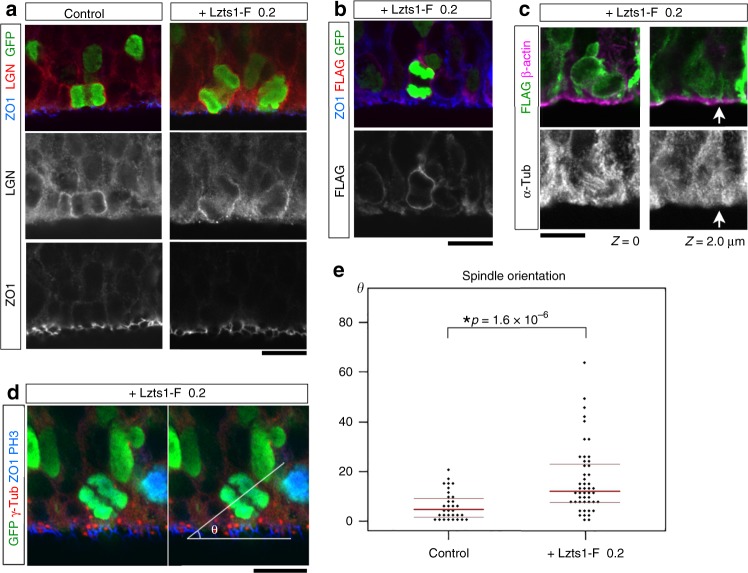
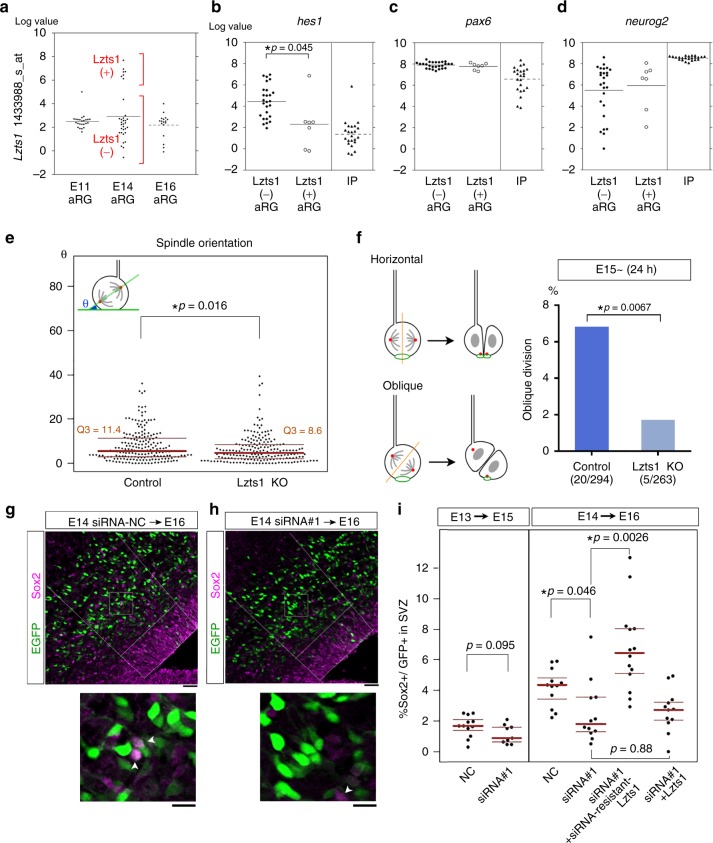
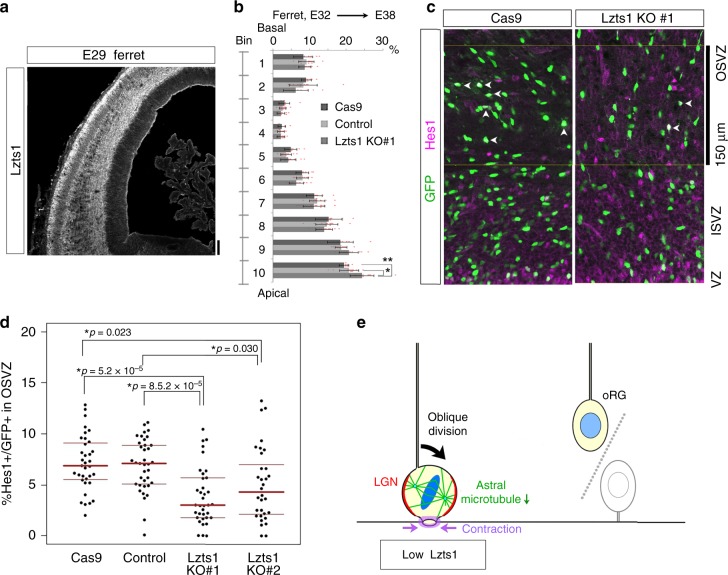
In the developing central nervous system, cell departure from the apical surface is the initial and fundamental step to form the 3D, organized architecture. Both delamination of differentiating cells and repositioning of progenitors to generate outer radial glial cells (oRGs) contribute to mammalian neocortical expansion; however, a comprehensive understanding of their mechanisms is lacking. Here, we demonstrate that Lzts1, a molecule associated with microtubule components, promotes both cell departure events. In neuronally committed cells, Lzts1 functions in apical delamination by altering apical junctional organization. In apical RGs (aRGs), Lzts1 expression is variable, depending on Hes1 expression levels. According to its differential levels, Lzts1 induces diverse RG behaviors: planar division, oblique divisions of aRGs that generate oRGs, and their mitotic somal translocation. Loss-of-function of lzts1 impairs all these cell departure processes. Thus, Lzts1 functions as a master modulator of cellular dynamics, contributing to increasing complexity of the cerebral architecture during evolution.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Neuronal Delamination and Outer Radial Glia Generation in Neocortical Development.Front Cell Dev Biol. 2021 Feb 5;8:623573. doi: 10.3389/fcell.2020.623573. eCollection 2020. Front Cell Dev Biol. 2021. PMID: 33614631 Free PMC article. Review.
-
Gliogenesis in the outer subventricular zone promotes enlargement and gyrification of the primate cerebrum.Proc Natl Acad Sci U S A. 2019 Apr 2;116(14):7089-7094. doi: 10.1073/pnas.1822169116. Epub 2019 Mar 20. Proc Natl Acad Sci U S A. 2019. PMID: 30894491 Free PMC article.
-
HoxB genes regulate neuronal delamination in the trunk neural tube by controlling the expression of Lzts1.Development. 2021 Feb 18;148(4):dev195404. doi: 10.1242/dev.195404. Development. 2021. PMID: 33472847
-
The expression profile of the tumor suppressor gene Lzts1 suggests a role in neuronal development.Dev Dyn. 2012 May;241(5):984-94. doi: 10.1002/dvdy.23777. Epub 2012 Mar 30. Dev Dyn. 2012. PMID: 22419569
-
Extracellular Control of Radial Glia Proliferation and Scaffolding During Cortical Development and Pathology.Front Cell Dev Biol. 2020 Oct 16;8:578341. doi: 10.3389/fcell.2020.578341. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33178693 Free PMC article. Review.
Cited by
-
Regulation of Cell Delamination During Cortical Neurodevelopment and Implication for Brain Disorders.Front Neurosci. 2022 Feb 23;16:824802. doi: 10.3389/fnins.2022.824802. eCollection 2022. Front Neurosci. 2022. PMID: 35281509 Free PMC article. Review.
-
Early spinal cord development: from neural tube formation to neurogenesis.Nat Rev Neurosci. 2025 Apr;26(4):195-213. doi: 10.1038/s41583-025-00906-5. Epub 2025 Feb 6. Nat Rev Neurosci. 2025. PMID: 39915695 Review.
-
Amelioration of biased neuronal differentiation in humanized mouse model of valproic acid-induced autism by precisely targeted transcranial magnetic stimulation.Bioeng Transl Med. 2025 Jan 23;10(3):e10748. doi: 10.1002/btm2.10748. eCollection 2025 May. Bioeng Transl Med. 2025. PMID: 40385547 Free PMC article.
-
Neuronal Delamination and Outer Radial Glia Generation in Neocortical Development.Front Cell Dev Biol. 2021 Feb 5;8:623573. doi: 10.3389/fcell.2020.623573. eCollection 2020. Front Cell Dev Biol. 2021. PMID: 33614631 Free PMC article. Review.
-
Advanced Techniques Using In Vivo Electroporation to Study the Molecular Mechanisms of Cerebral Development Disorders.Int J Mol Sci. 2023 Sep 15;24(18):14128. doi: 10.3390/ijms241814128. Int J Mol Sci. 2023. PMID: 37762431 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
