

Inhibition of hepatocellular carcinoma by metabolic normalization
- PMID: 31242205
- PMCID: PMC6594671
- DOI: 10.1371/journal.pone.0218186
Inhibition of hepatocellular carcinoma by metabolic normalization
Abstract
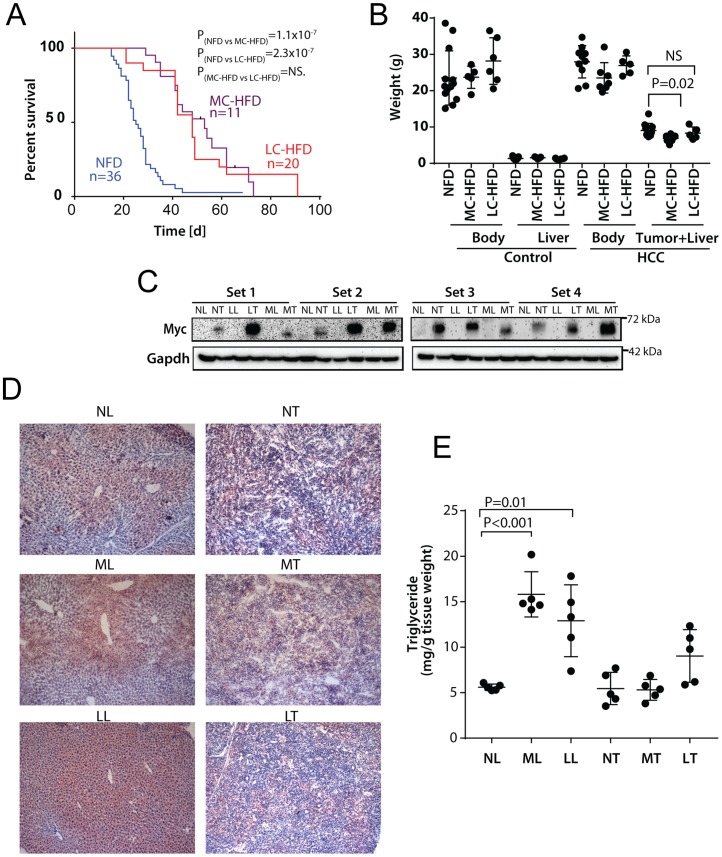
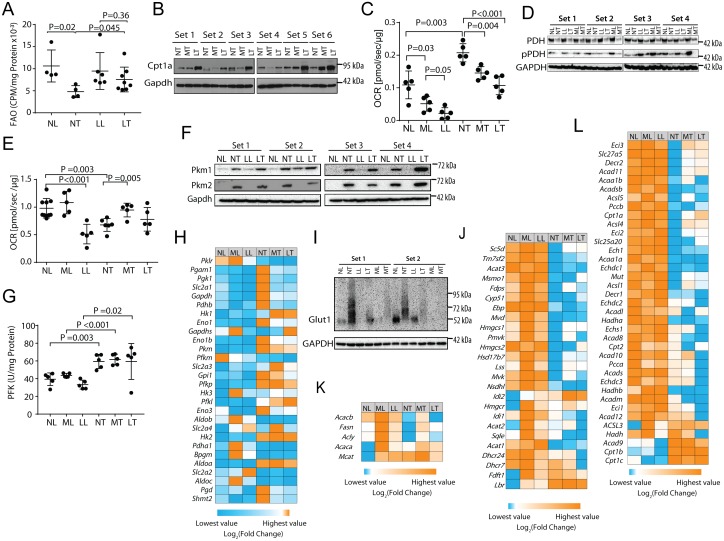
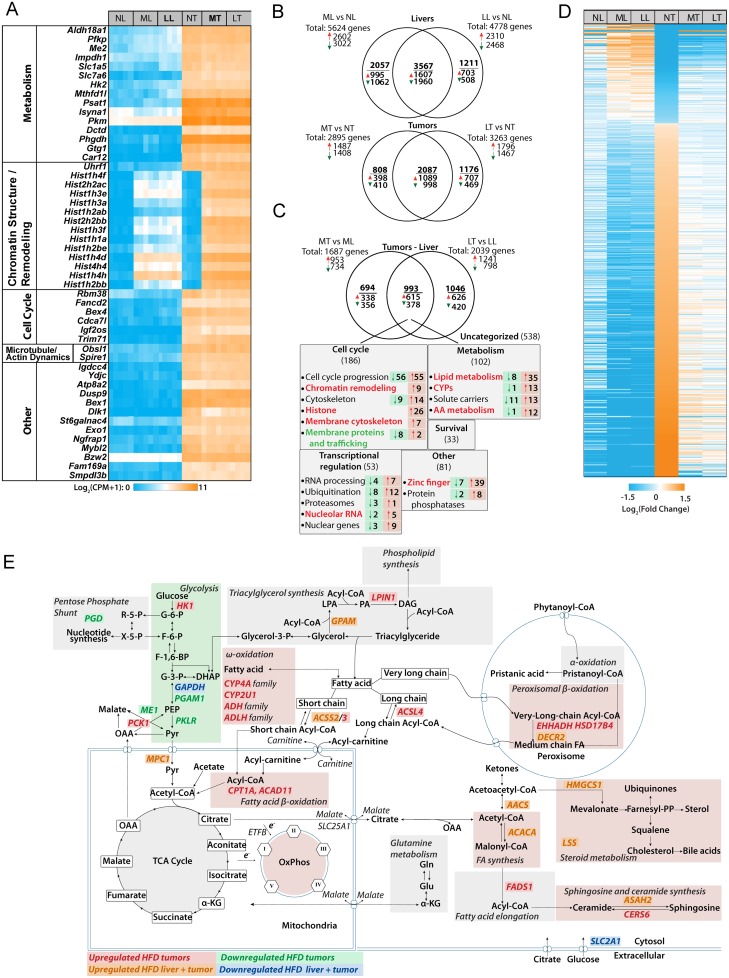
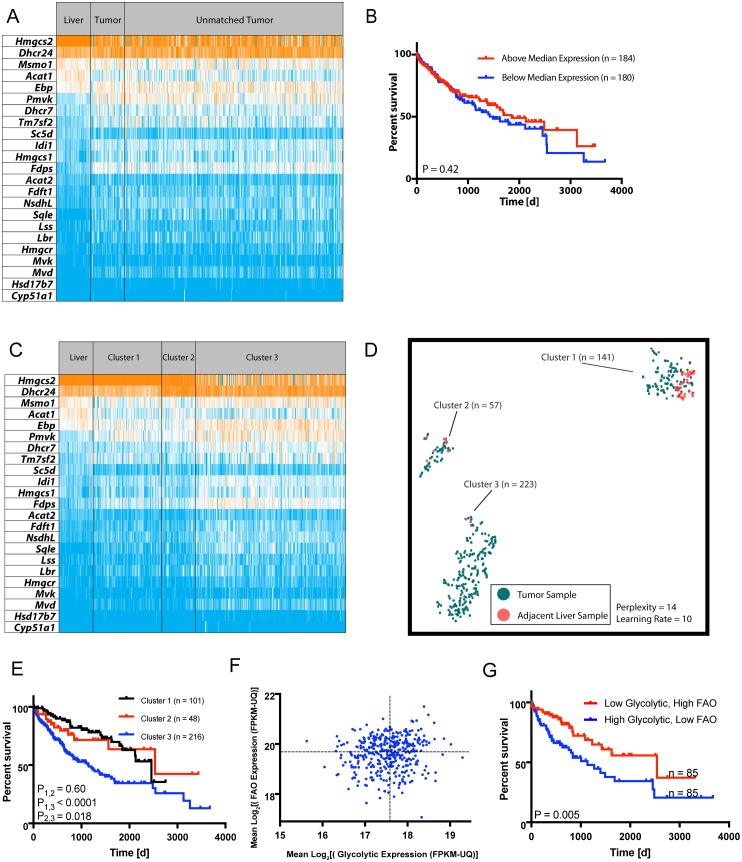
In two different mouse liver cancer models, we recently showed that a switch from oxidative phosphorylation (Oxphos) to glycolysis (the Warburg effect) is invariably accompanied by a marked decline in fatty acid oxidation (FAO) and a reciprocal increase in the activity of pyruvate dehydrogenase (PDH), which links glycolysis to the TCA cycle. We now show that short-term implementation of either medium-chain (MC) or long-chain (LC) high fat diets (HFDs) nearly doubled the survival of mice with c-Myc oncoprotein-driven hepatocellular carcinoma (HCC). Mechanistically, HFDs forced tumors to become more reliant on fatty acids as an energy source, thus normalizing both FAO and PDH activities. More generally, both MC- and LC-HFDs partially or completely normalized the expression of 682 tumor-dysregulated transcripts, a substantial fraction of which are involved in cell cycle control, proliferation and metabolism. That these same transcripts were responsive to HFDs in livers strongly suggested that the changes were the cause of tumor inhibition rather than its consequence. In seven different human cancer cohorts, patients with tumors containing high ratios of FAO-related:glycolysis-related transcripts had prolonged survival relative to those with low ratios. Furthermore, in 13 human cancer types, the expression patterns of transcripts encoding enzymes participating in FAO and/or cholesterol biosynthesis also correlated with significantly prolonged survival. Collectively, our results support the idea that the survival benefits of HFDs are due to a reversal of the Warburg effect and other tumor-associated metabolic and cell cycle abnormalities. They also suggest that short-term dietary manipulation, either alone or in combination with more traditional chemotherapeutic regimens, might be employed as a relatively non-toxic and cost-effective means of enhancing survival in certain cancer types.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
