

The protective effect of inflammatory monocytes during systemic C. albicans infection is dependent on collaboration between C-type lectin-like receptors
- PMID: 31242262
- PMCID: PMC6594653
- DOI: 10.1371/journal.ppat.1007850
The protective effect of inflammatory monocytes during systemic C. albicans infection is dependent on collaboration between C-type lectin-like receptors
Abstract
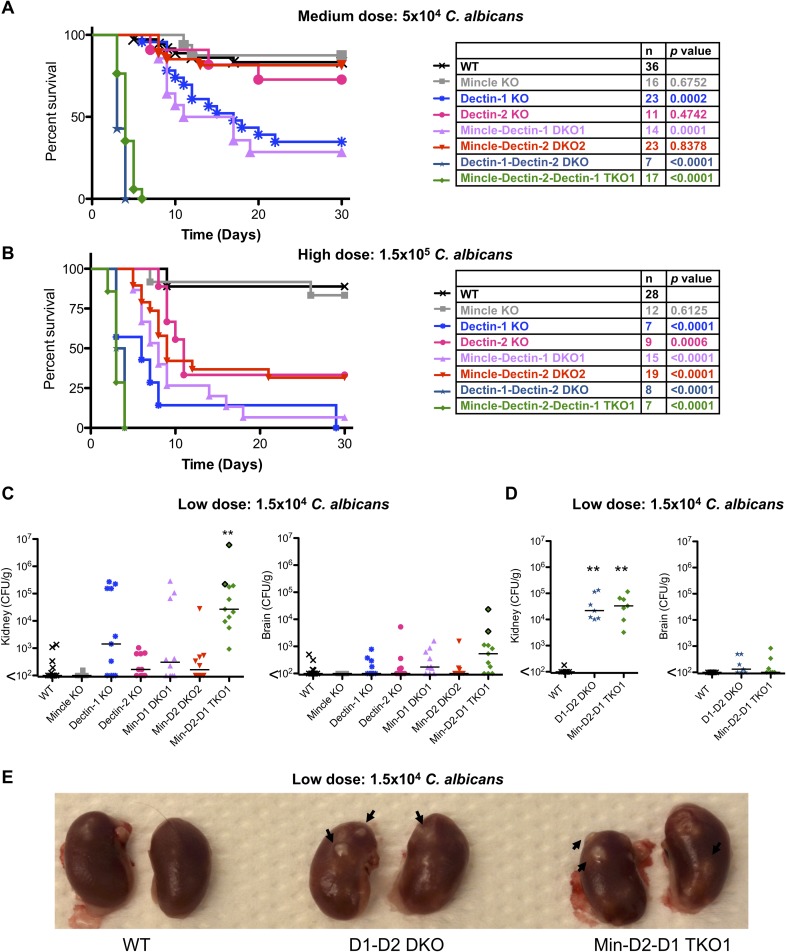
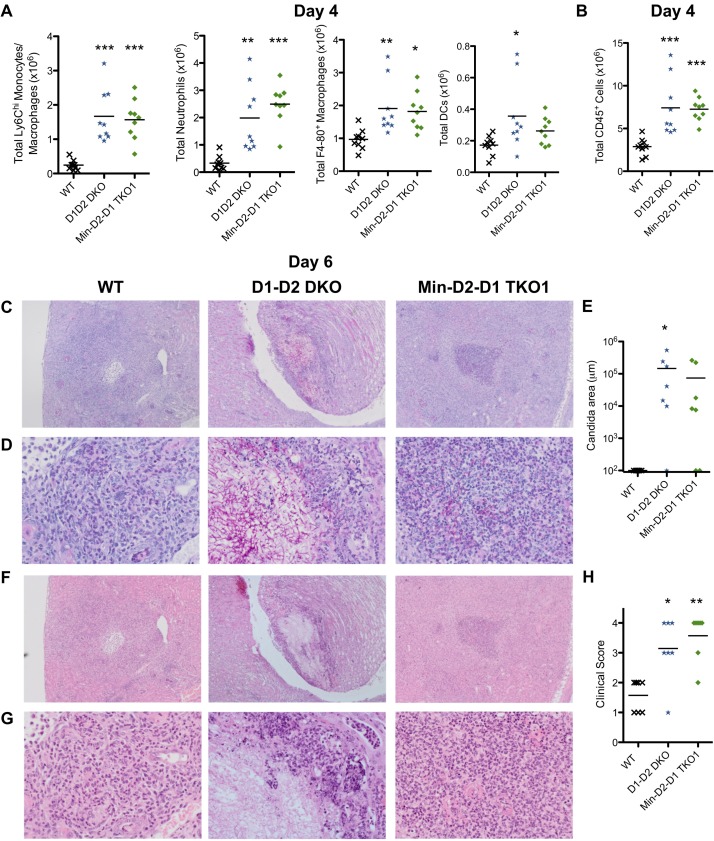
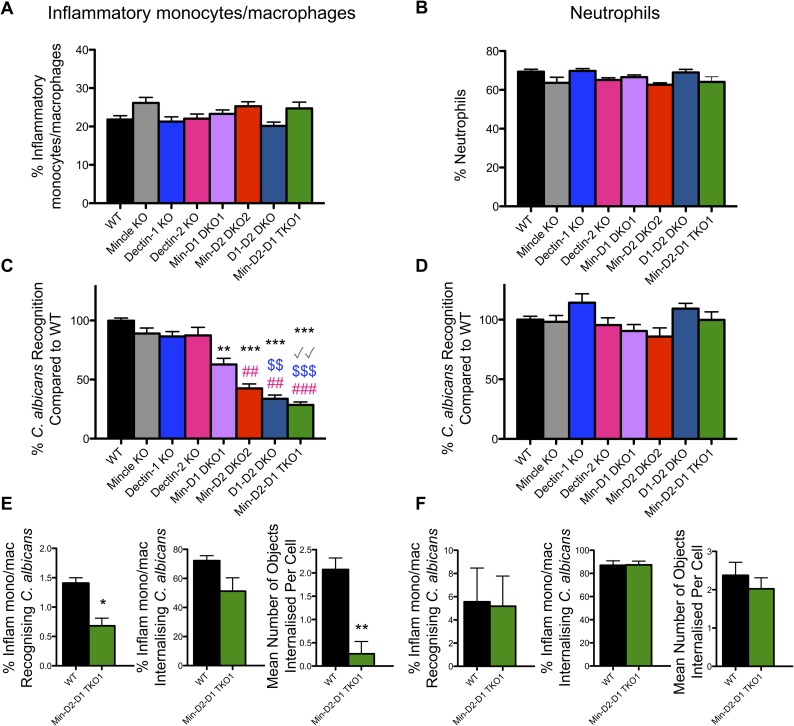
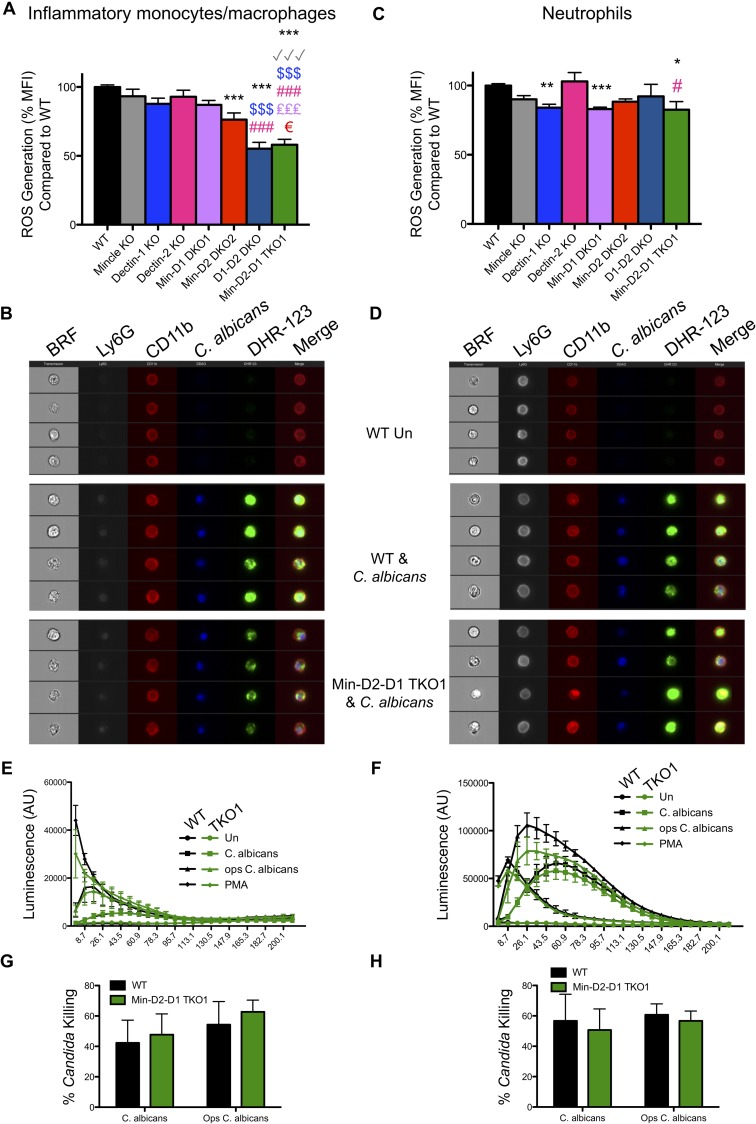
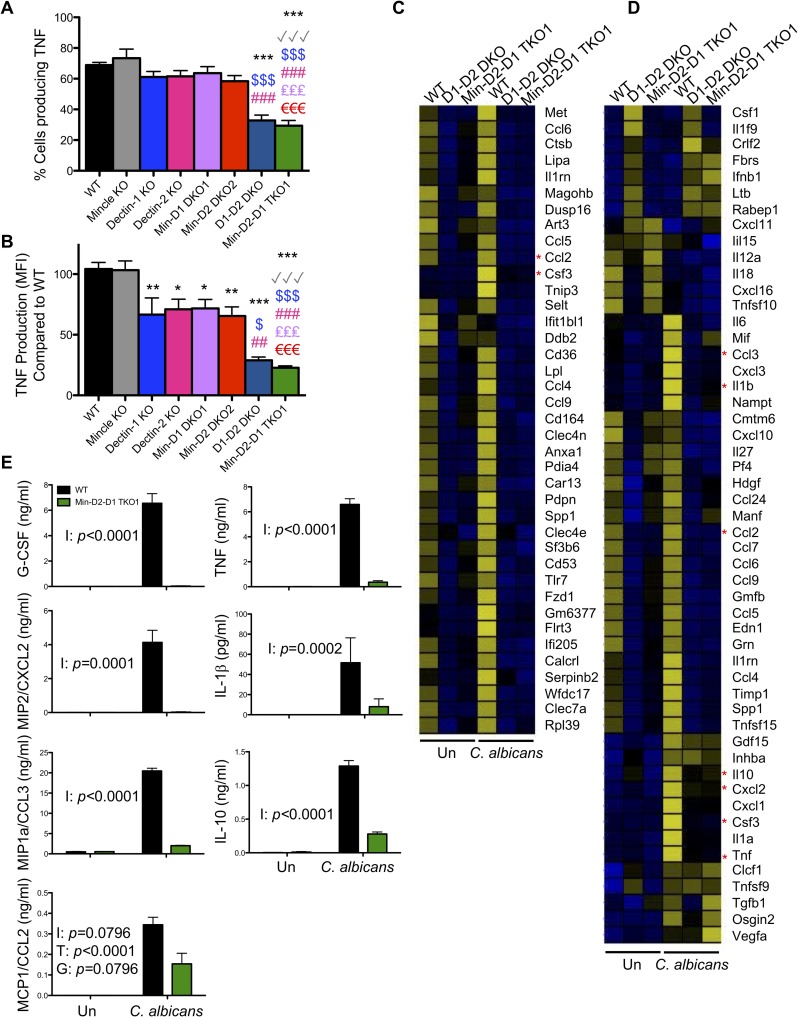
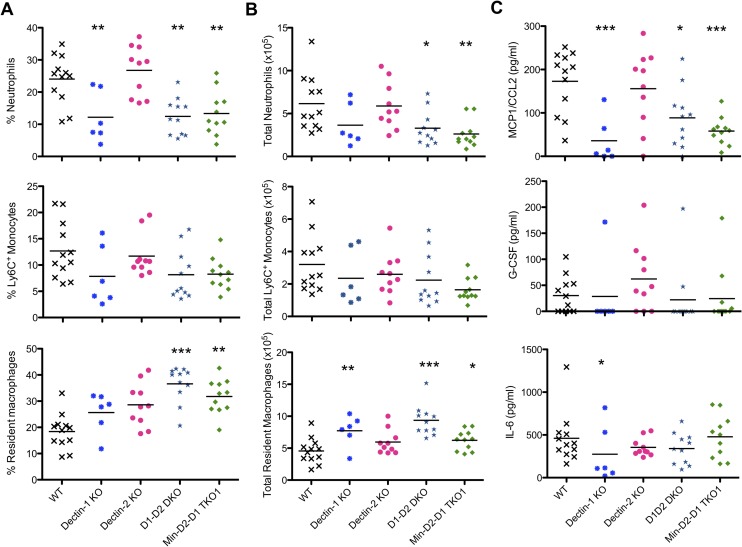
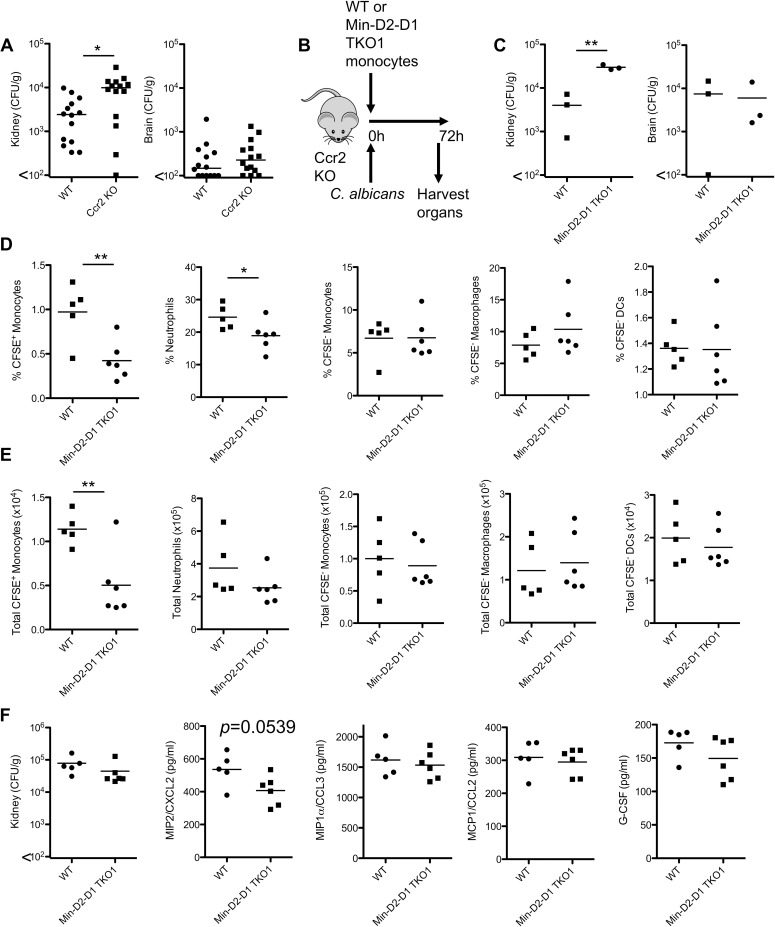
Invasive candidiasis, mainly caused by Candida albicans, is a serious healthcare problem with high mortality rates, particularly in immunocompromised patients. Innate immune cells express pathogen recognition receptors (PRRs) including C-type lectin-like receptors (CLRs) that bind C. albicans to initiate an immune response. Multiple CLRs including Dectin-1, Dectin-2 and Mincle have been proposed individually to contribute to the immune response to C. albicans. However how these receptors collaborate to clear a fungal infection is unknown. Herein, we used novel multi-CLR knockout (KO) mice to decipher the individual, collaborative and collective roles of Dectin-1, Dectin-2 and Mincle during systemic C. albicans infection. These studies revealed an unappreciated and profound role for CLR co-operation in anti-fungal immunity. The protective effect of multiple CLRs was markedly greater than any single receptor, and was mediated through inflammatory monocytes via recognition and phagocytosis of C. albicans, and production of C. albicans-induced cytokines and chemokines. These CLRs were dispensable for mediating similar responses from neutrophils, likely due to lower expression of these CLRs on neutrophils compared to inflammatory monocytes. Concurrent deletion of Dectin-1 and Dectin-2, or all three CLRs, resulted in dramatically increased susceptibility to systemic C. albicans infection compared to mice lacking a single CLR. Multi-CLR KO mice were unable to control fungal growth due to an inadequate early inflammatory monocyte-mediated response. In response to excessive fungal growth, the multi-CLR KO mice mounted a hyper-inflammatory response, likely leading to multiple organ failure. Thus, these data reveal a critical role for CLR co-operation in the effective control of C. albicans and maintenance of organ function during infection.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Strasser D, Neumann K, Bergmann H, Marakalala MJ, Guler R, Rojowska A, et al. Syk kinase-coupled C-type lectin receptors engage protein kinase C-sigma to elicit Card9 adaptor-mediated innate immunity. Immunity. 2012;36(1):32–42. Epub 2012/01/24. S1074-7613(12)00002-7 [pii] 10.1016/j.immuni.2011.11.015 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
