

Early Seizure Activity Accelerates Depletion of Hippocampal Neural Stem Cells and Impairs Spatial Discrimination in an Alzheimer's Disease Model
- PMID: 31242408
- PMCID: PMC6697001
- DOI: 10.1016/j.celrep.2019.05.101
Early Seizure Activity Accelerates Depletion of Hippocampal Neural Stem Cells and Impairs Spatial Discrimination in an Alzheimer's Disease Model
Abstract
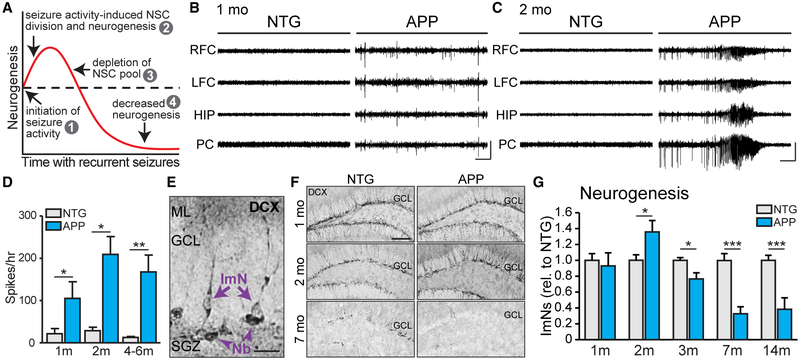
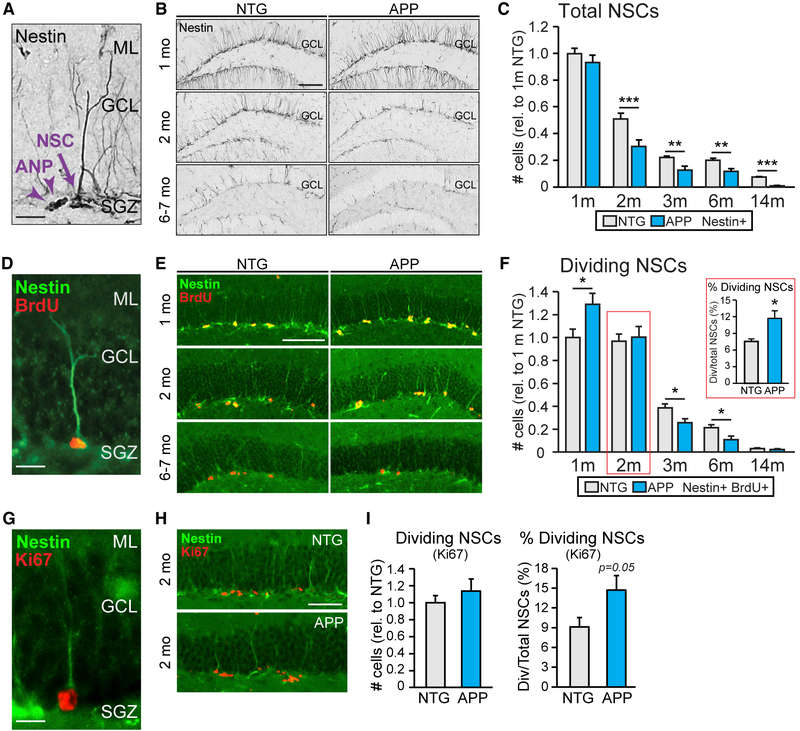
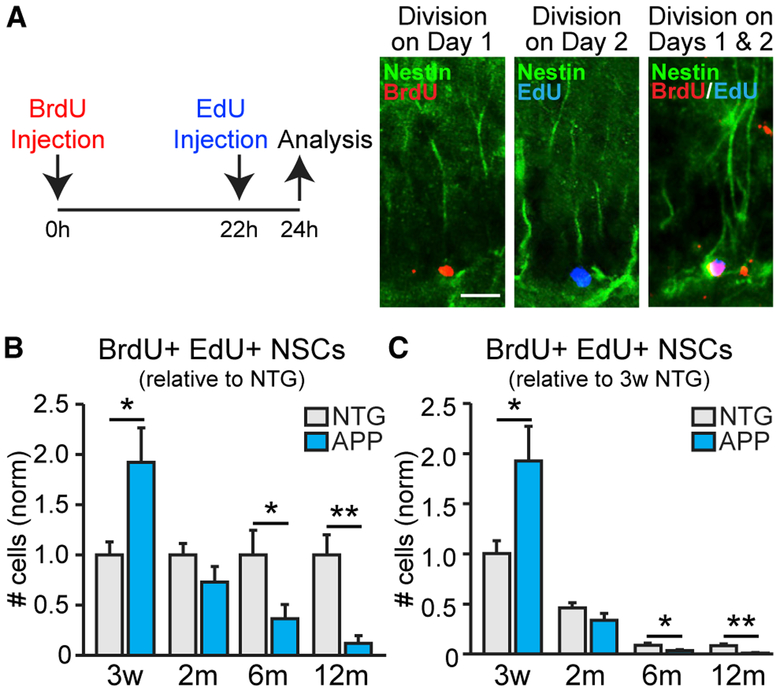
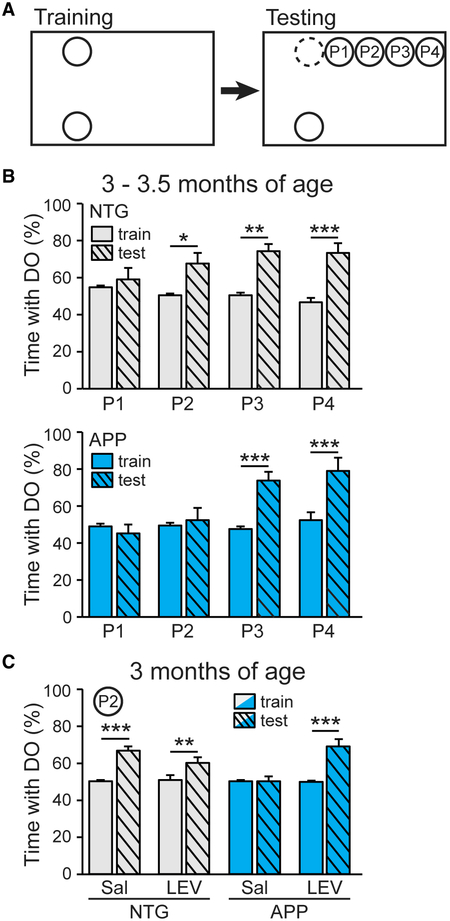
Adult hippocampal neurogenesis has been reported to be decreased, increased, or not changed in Alzheimer's disease (AD) patients and related transgenic mouse models. These disparate findings may relate to differences in disease stage, or the presence of seizures, which are associated with AD and can stimulate neurogenesis. In this study, we investigate a transgenic mouse model of AD that exhibits seizures similarly to AD patients and find that neurogenesis is increased in early stages of disease, as spontaneous seizures became evident, but is decreased below control levels as seizures recur. Treatment with the antiseizure drug levetiracetam restores neurogenesis and improves performance in a neurogenesis-associated spatial discrimination task. Our results suggest that seizures stimulate, and later accelerate the depletion of, the hippocampal neural stem cell pool. These results have implications for AD as well as any disorder accompanied by recurrent seizures, such as epilepsy.
Keywords: Alzheimer; cognition; dentate gyrus; epilepsy; hippocampus; memory; mouse model; neural stem cell pool; neurogenesis; seizure.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures

References
-
- Apostolopoulou M, Kiehl TR, Winter M, Cardenas De La Hoz E, Boles NC, Bjornsson CS, Zuloaga KL, Goderie SK, Wang Y, Cohen AR, and Temple S (2017). Non-monotonic Changes in Progenitor Cell Behavior and Gene Expression during Aging of the Adult V-SVZ Neural Stem Cell Niche. Stem Cell Rep. 9, 1931–1947. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
