

CRISPR-READI: Efficient Generation of Knockin Mice by CRISPR RNP Electroporation and AAV Donor Infection
- PMID: 31242412
- PMCID: PMC6693498
- DOI: 10.1016/j.celrep.2019.05.103
CRISPR-READI: Efficient Generation of Knockin Mice by CRISPR RNP Electroporation and AAV Donor Infection
Abstract
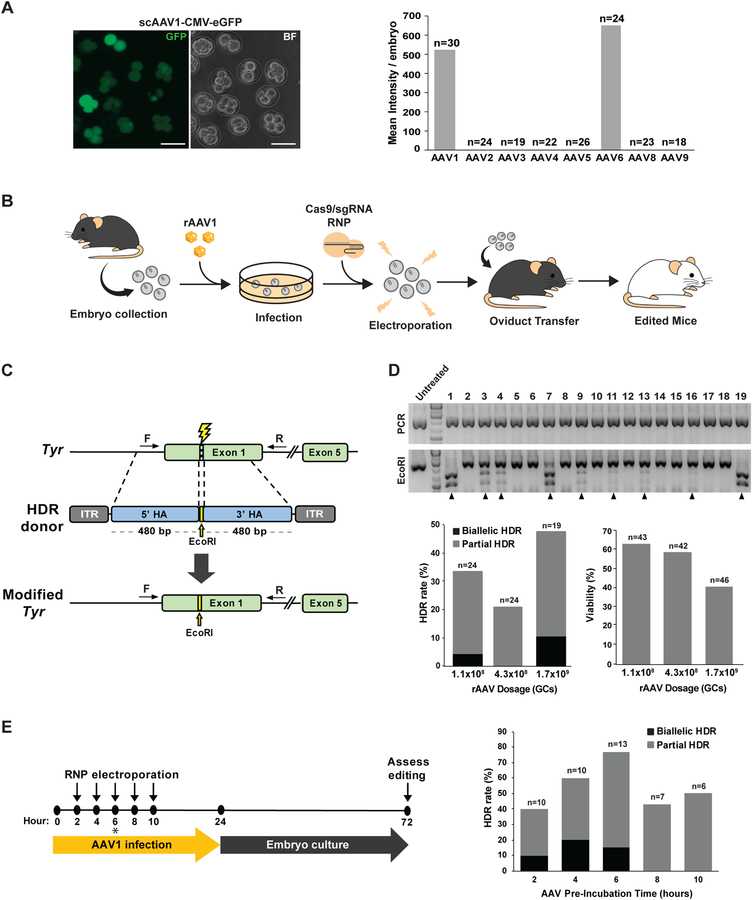
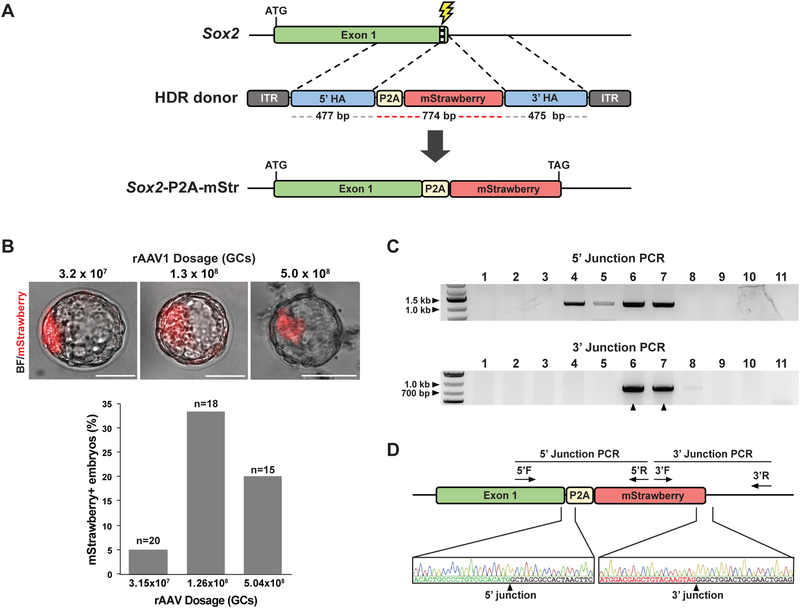
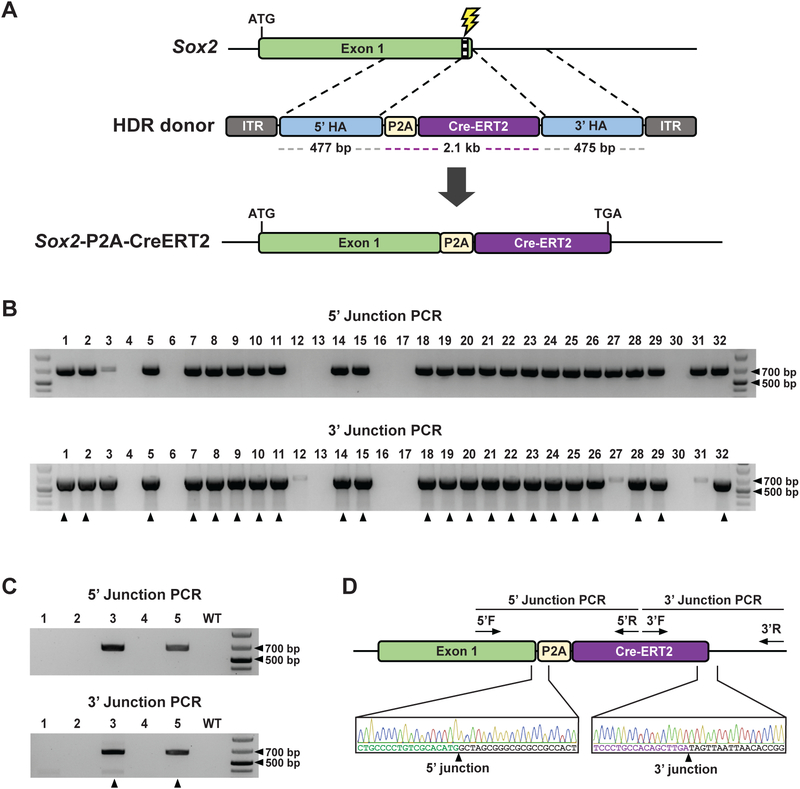
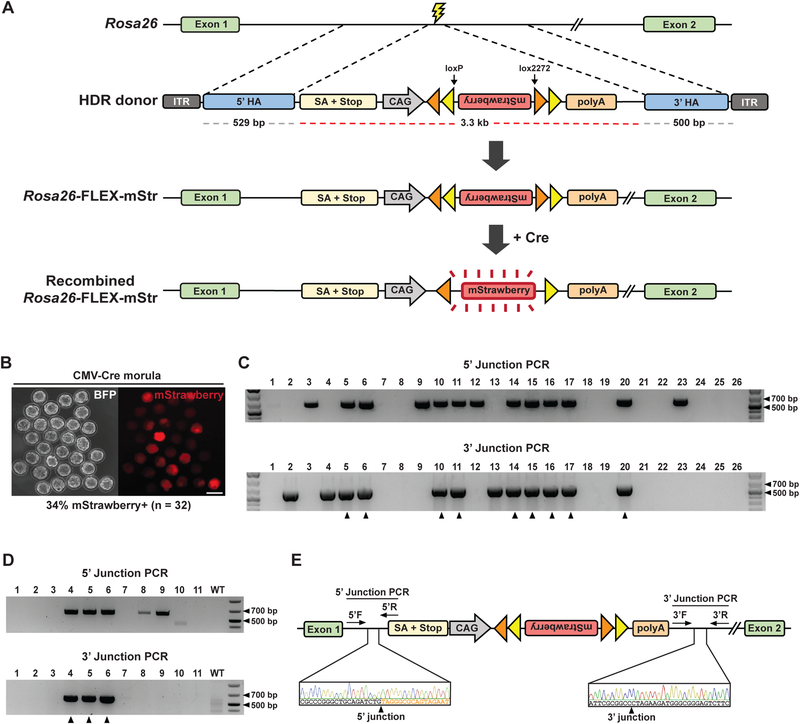
Genetically engineered mouse models harboring large sequence insertions or modifications are critical for a wide range of applications including endogenous gene tagging, conditional knockout, site-specific transgene insertion, and gene replacement; however, existing methods to generate such animals remain laborious and costly. To address this, we developed an approach called CRISPR-READI (CRISPR RNP electroporation and AAV donor infection), combining adeno-associated virus (AAV)-mediated HDR donor delivery with Cas9/sgRNA RNP electroporation to engineer large site-specific modifications in the mouse genome with high efficiency and throughput. We successfully targeted a 774 bp fluorescent reporter, a 2.1 kb CreERT2 driver, and a 3.3 kb expression cassette into endogenous loci in both embryos and live mice. CRISPR-READI is applicable to most widely used knockin schemes requiring donor lengths within the 4.9 kb AAV packaging capacity. Altogether, CRISPR-READI is an efficient, high-throughput, microinjection-free approach for sophisticated mouse genome engineering with potential applications in other mammalian species.
Keywords: AAV; CRISPR; CRISPR-EZ; HDR editing; electroporation; genome editing; knockin; mouse models.
Copyright © 2019. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests
DVS is an inventor on patents involving AAV directed evolution and a co-founder and a shareholder of a company developing AAV vectors for clinical gene therapy. In addition, he is on the board of directors of and holds shares in a second company developing AAV vectors for clinical gene therapy. SC and LH are inventors on patents involving an electroporation-based CRISPR technology for mouse genome engineering and are founders of a company to further develop this technology for mammalian genome editing.
Figures
References
-
- Bao X, Lian X, Hacker TA, Schmuck EG, Qian T, Bhute VJ, Han T, Shi M, Drowley L, Plowright A, Wang Q-D, Goumans M-J, Palecek SP, 2016. Long-term self-renewing human epicardial cells generated from pluripotent stem cells under defined xeno-free conditions. Nat. Biomed. Eng 1 10.1038/s41551-016-0003 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
