

Pediatric patients with acute lymphoblastic leukemia generate abundant and functional neoantigen-specific CD8+ T cell responses
- PMID: 31243155
- PMCID: PMC7020562
- DOI: 10.1126/scitranslmed.aat8549
Pediatric patients with acute lymphoblastic leukemia generate abundant and functional neoantigen-specific CD8+ T cell responses
Abstract
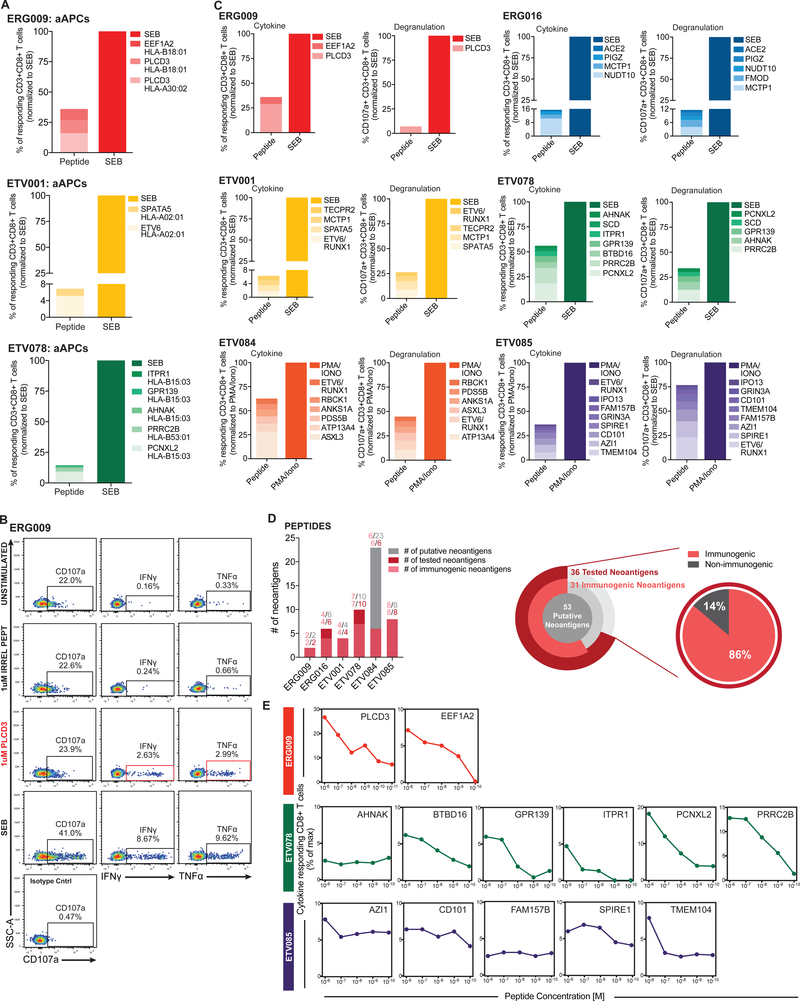
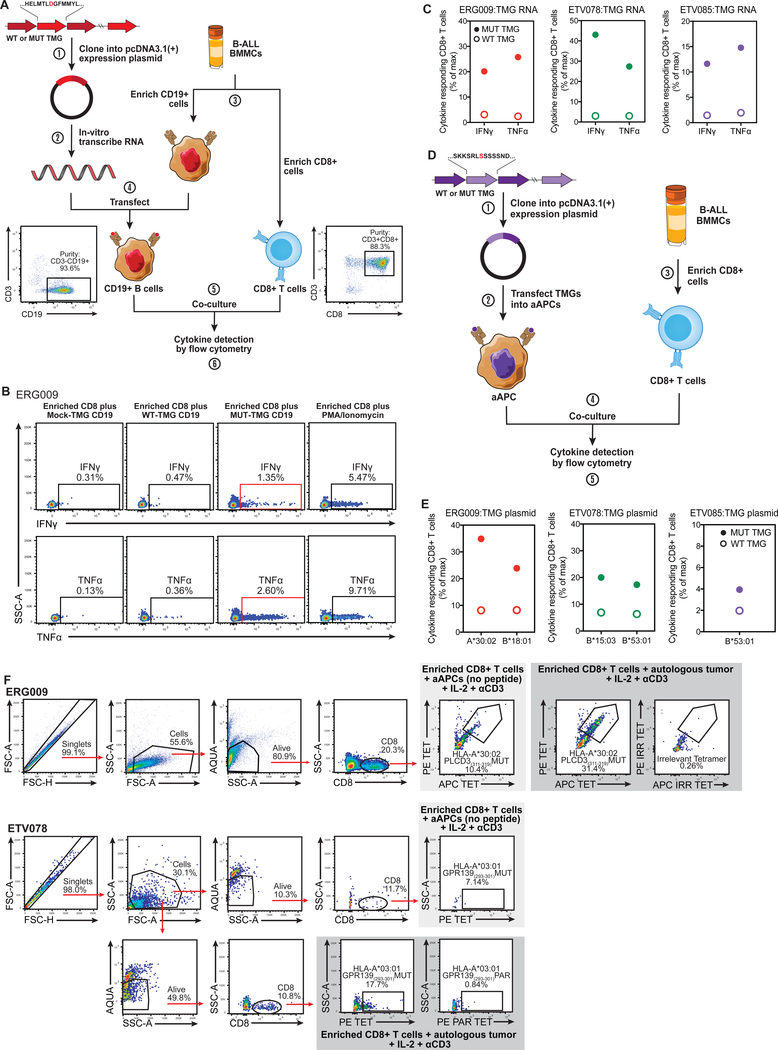
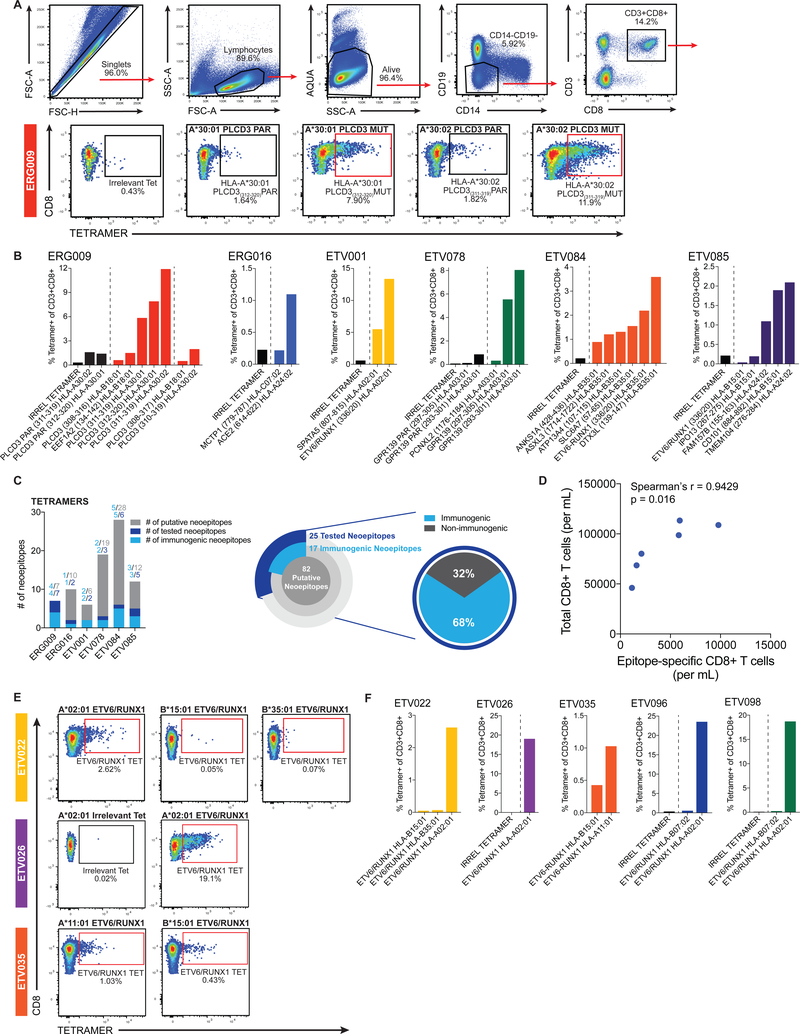
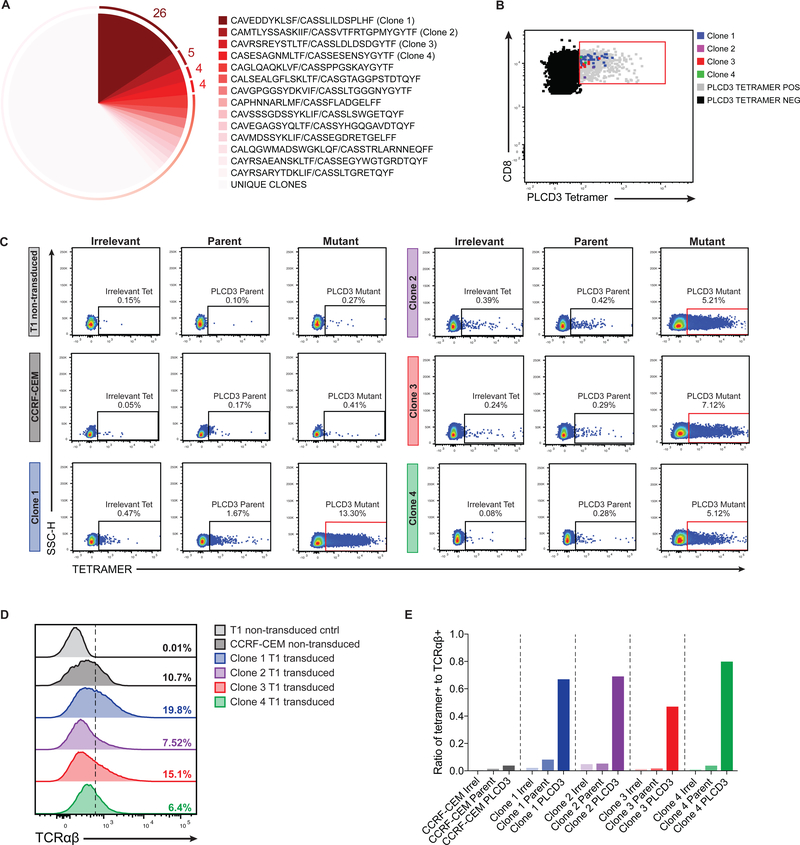
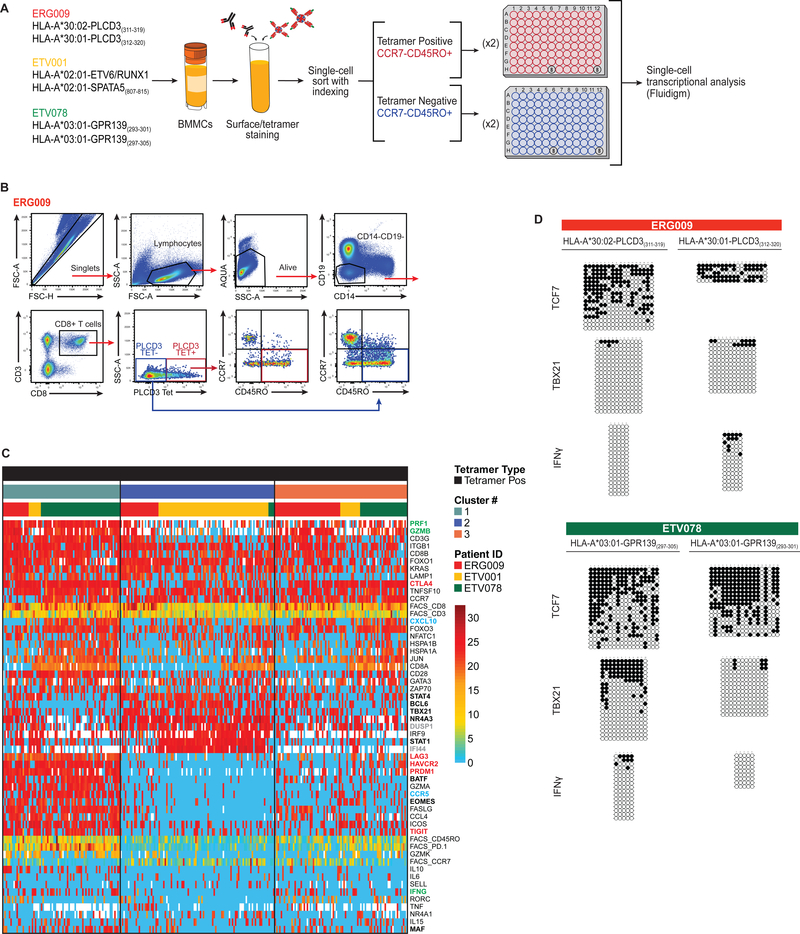
Cancer arises from the accumulation of genetic alterations, which can lead to the production of mutant proteins not expressed by normal cells. These mutant proteins can be processed and presented on the cell surface by major histocompatibility complex molecules as neoepitopes, allowing CD8+ T cells to mount responses against them. For solid tumors, only an average 2% of neoepitopes predicted by algorithms have detectable endogenous antitumor T cell responses. This suggests that low mutation burden tumors, which include many pediatric tumors, are poorly immunogenic. Here, we report that pediatric patients with acute lymphoblastic leukemia (ALL) have tumor-associated neoepitope-specific CD8+ T cells, responding to 86% of tested neoantigens and recognizing 68% of the tested neoepitopes. These responses include a public neoantigen from the ETV6-RUNX1 fusion that is targeted in seven of nine tested patients. We characterized phenotypic and transcriptional profiles of CD8+ tumor-infiltrating lymphocytes (TILs) at the single-cell level and found a heterogeneous population that included highly functional effectors. Moreover, we observed immunodominance hierarchies among the CD8+ TILs restricted to one or two putative neoepitopes. Our results indicate that robust antitumor immune responses are induced in pediatric ALL despite their low mutation burdens and emphasize the importance of immunodominance in shaping cellular immune responses. Furthermore, these data suggest that pediatric cancers may be amenable to immunotherapies aimed at enhancing immune recognition of tumor-specific neoantigens.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Challenging (immunogenically) silent assumptions.Nat Rev Cancer. 2019 Sep;19(9):483. doi: 10.1038/s41568-019-0181-1. Nat Rev Cancer. 2019. PMID: 31316158 No abstract available.
-
Childhood leukemia could be a target for immunotherapy.Cancer. 2019 Dec 15;125(24):4345. doi: 10.1002/cncr.32633. Cancer. 2019. PMID: 31792938 No abstract available.
References
-
- Schumacher TN, Schreiber RD, Neoantigens in cancer immunotherapy, Science 348, 69–74 (2015). - PubMed
-
- Robbins PF, Kassim SH, Tran TLN, Crystal JS, Morgan RA, Feldman SA, Yang JC, Dudley ME, Wunderlich JR, Sherry RM, Kammula US, Hughes MS, Restifo NP, Raffeld M, Lee C-CR, Li YF, El-Gamil M, Rosenberg SA, A Pilot Trial Using Lymphocytes Genetically Engineered with an NY-ESO-1–Reactive T-cell Receptor: Long-term Follow-up and Correlates with Response, Clin. Cancer Res. 21, 1019–1027 (2015). - PMC - PubMed
-
- Strønen E, Toebes M, Kelderman S, van Buuren MM, Yang W, van Rooij N, Donia M, Böschen M-L, Lund-Johansen F, Olweus J, Schumacher TN, Targeting of cancer neoantigens with donor-derived T cell receptor repertoires, Science 352, 1337–1341 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
