

Comment on 'AIRE-deficient patients harbor unique high-affinity disease-ameliorating autoantibodies'
- PMID: 31244471
- PMCID: PMC6597240
- DOI: 10.7554/eLife.43578
Comment on 'AIRE-deficient patients harbor unique high-affinity disease-ameliorating autoantibodies'
Abstract
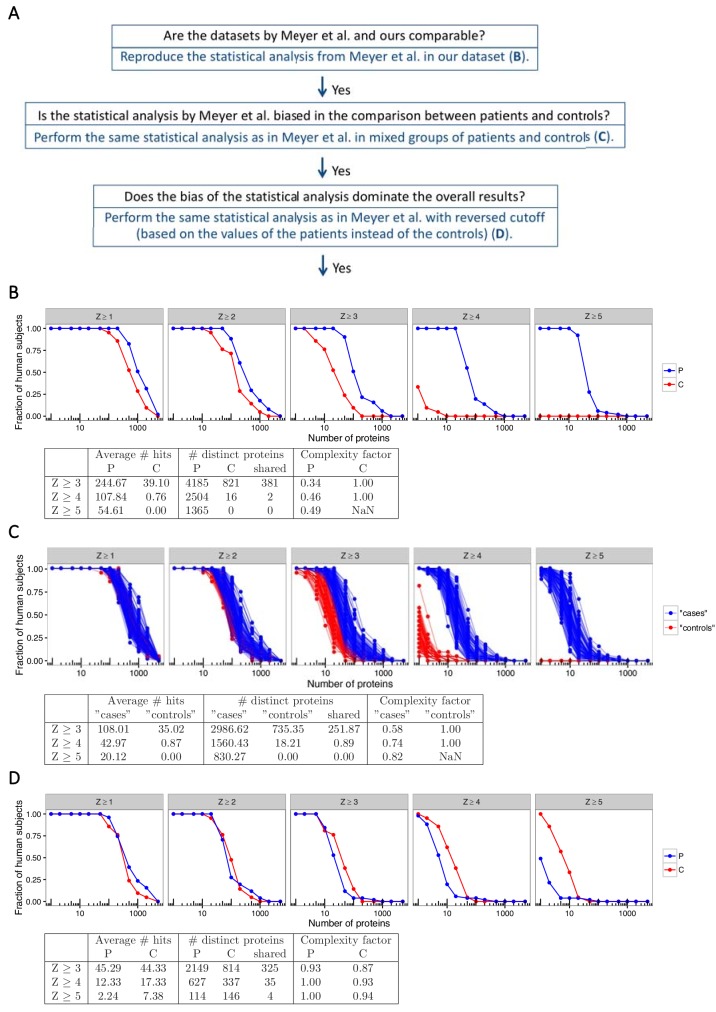
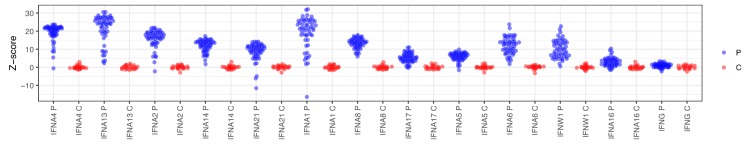
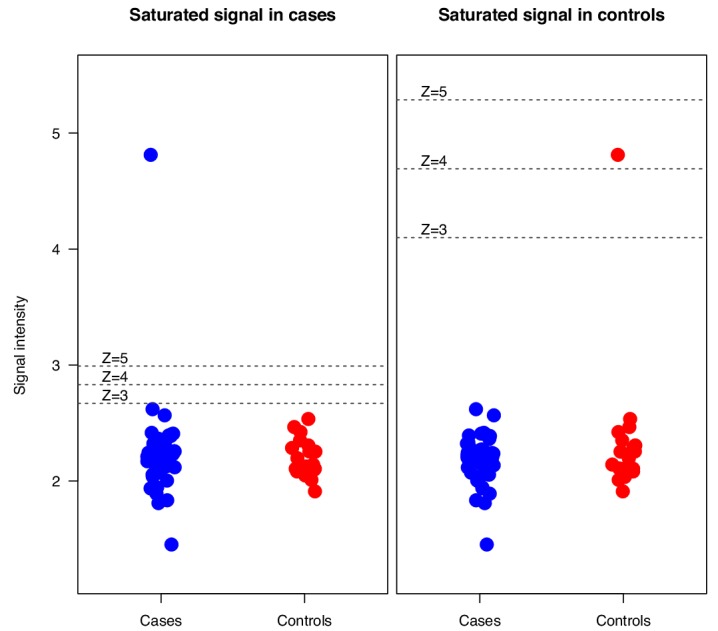
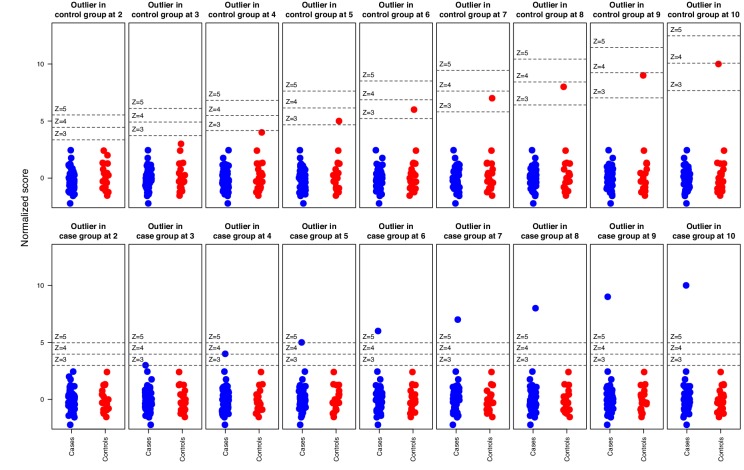
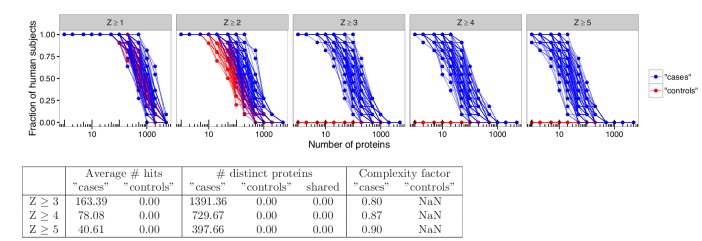
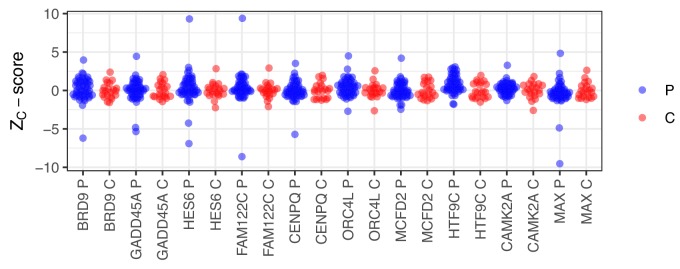
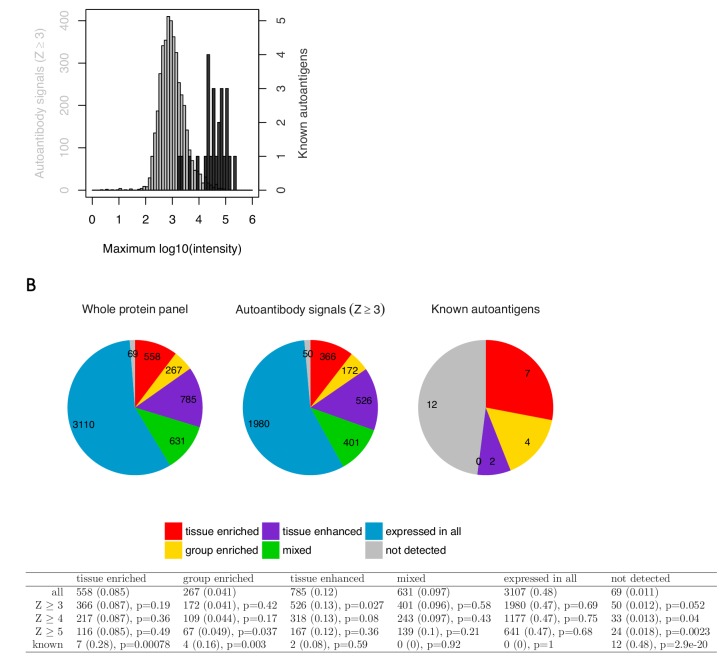
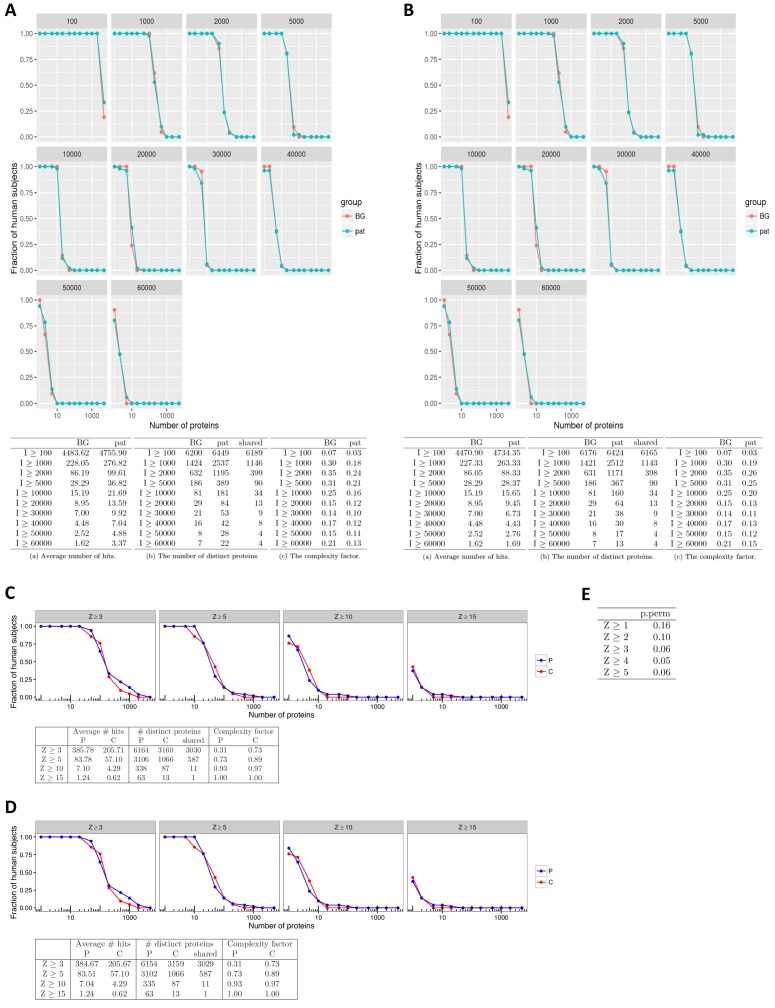
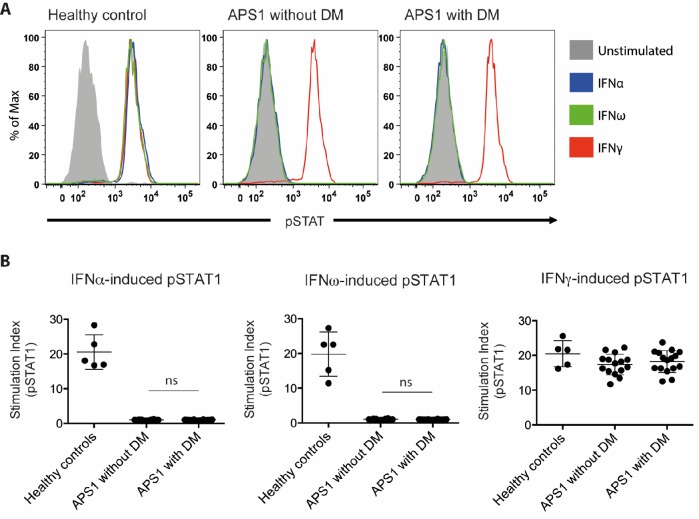
The AIRE gene plays a key role in the development of central immune tolerance by promoting thymic presentation of tissue-specific molecules. Patients with AIRE-deficiency develop multiple autoimmune manifestations and display autoantibodies against the affected tissues. In 2016 it was reported that: i) the spectrum of autoantibodies in patients with AIRE-deficiency is much broader than previously appreciated; ii) neutralizing autoantibodies to type I interferons (IFNs) could provide protection against type 1 diabetes in these patients (Meyer et al., 2016). We attempted to replicate these new findings using a similar experimental approach in an independent patient cohort, and found no evidence for either conclusion.
Keywords: APS1/APECED; autoantibody; autoantigen; human; human biology; immune tolerance; immunology; inflammation; medicine; type 1 diabetes.
Conflict of interest statement
NL, LR, EF, DE, TF, GS, EF, PB, DS, ML, MA No competing interests declared, MS Serves as founder and consultant for Personalis, is a member of the scientific advisory board of GenapSys, and a consultant for Illumina. OK Is a board member of Olink Bioscience.
Figures
Comment in
-
Response to comment on 'AIRE-deficient patients harbor unique high-affinity disease-ameliorating autoantibodies'.Elife. 2019 Jun 27;8:e45826. doi: 10.7554/eLife.45826. Elife. 2019. PMID: 31244472 Free PMC article.
Comment on
-
AIRE-Deficient Patients Harbor Unique High-Affinity Disease-Ameliorating Autoantibodies.Cell. 2016 Jul 28;166(3):582-595. doi: 10.1016/j.cell.2016.06.024. Epub 2016 Jul 14. Cell. 2016. PMID: 27426947 Free PMC article.
References
-
- Alimohammadi M, Björklund P, Hallgren A, Pöntynen N, Szinnai G, Shikama N, Keller MP, Ekwall O, Kinkel SA, Husebye ES, Gustafsson J, Rorsman F, Peltonen L, Betterle C, Perheentupa J, Akerström G, Westin G, Scott HS, Holländer GA, Kämpe O. Autoimmune polyendocrine syndrome type 1 and NALP5, a parathyroid autoantigen. New England Journal of Medicine. 2008;358:1018–1028. doi: 10.1056/NEJMoa0706487. - DOI - PubMed
-
- Alimohammadi M, Dubois N, Sköldberg F, Hallgren A, Tardivel I, Hedstrand H, Haavik J, Husebye ES, Gustafsson J, Rorsman F, Meloni A, Janson C, Vialettes B, Kajosaari M, Egner W, Sargur R, Pontén F, Amoura Z, Grimfeld A, De Luca F, Betterle C, Perheentupa J, Kämpe O, Carel JC. Pulmonary autoimmunity as a feature of autoimmune polyendocrine syndrome type 1 and identification of KCNRG as a bronchial autoantigen. PNAS. 2009;106:4396–4401. doi: 10.1073/pnas.0809986106. - DOI - PMC - PubMed
