

Spring flowering habit in field pennycress (Thlaspi arvense) has arisen multiple independent times
- PMID: 31245698
- PMCID: PMC6508777
- DOI: 10.1002/pld3.97
Spring flowering habit in field pennycress (Thlaspi arvense) has arisen multiple independent times
Abstract
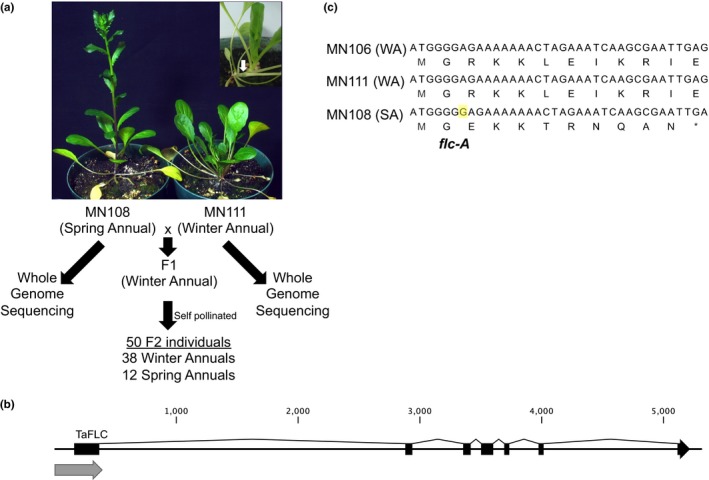
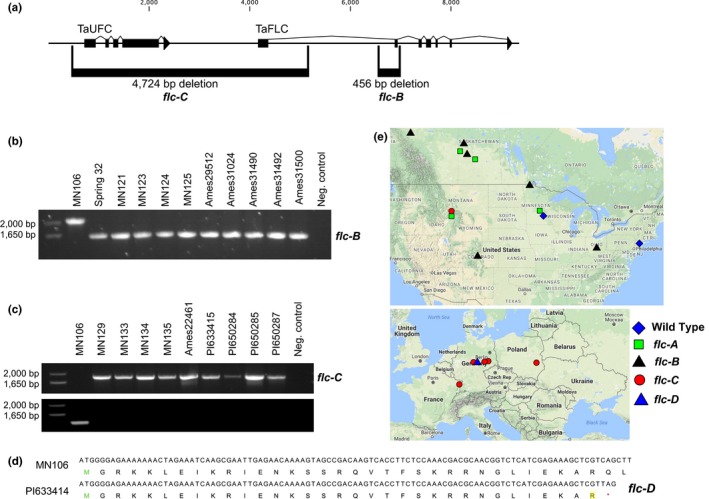
Field pennycress (Thlaspi arvense L.) is currently being developed as a new cold-tolerant oilseed crop. In natural populations, pennycress, like many Brassicaceae relatives, can exhibit either a winter or spring annual phenotype. Pennycress is a diploid relative of Arabidopsis thaliana, a model species that has been used to study many adaptive phenotypes, including flowering time and developmental timing. In Arabidopsis and other Brassicaceae species, mutations in negative regulators of flowering, including FLOWERING LOCUS C and FRIGIDA can cause the transition to a spring annual habit. The genetics underlying the difference between spring and winter annual pennycress lines are currently unknown. Here, we report the identification of four natural alleles of FLC in pennycress that confer a spring annual growth habit identified through whole genome sequencing, cosegregation analyses, and comparative genomics. The global distribution of these spring annual alleles of FLC suggests that the spring annual growth habit has arisen on several independent occasions. The two spring annual FLC alleles present in European accessions were only identified in North American accessions collected in southern Montana, which indicates accessions harboring these two alleles were introduced to North America, likely after pennycress became a widespread species on the continent. These findings provide new information on the natural history of the introduction and spread of spring annual pennycress accessions from Europe into North America. At the molecular level, these findings are important for the ongoing development of pennycress as a winter annual crop. An enhanced understanding of the regulation of flowering in this species should allow for the fine-tuning of flowering in commercial varieties.
Keywords: Thlaspi arvense; flowering locus C; pennycress; spring annual; whole genome sequencing; winter annual.
Figures
Similar articles
-
Genomic analysis of field pennycress (Thlaspi arvense) provides insights into mechanisms of adaptation to high elevation.BMC Biol. 2021 Jul 22;19(1):143. doi: 10.1186/s12915-021-01079-0. BMC Biol. 2021. PMID: 34294107 Free PMC article.
-
New approaches to facilitate rapid domestication of a wild plant to an oilseed crop: example pennycress (Thlaspi arvense L.).Plant Sci. 2014 Oct;227:122-32. doi: 10.1016/j.plantsci.2014.07.008. Epub 2014 Aug 4. Plant Sci. 2014. PMID: 25219314 Review.
-
De novo assembly of the pennycress (Thlaspi arvense) transcriptome provides tools for the development of a winter cover crop and biodiesel feedstock.Plant J. 2013 Sep;75(6):1028-38. doi: 10.1111/tpj.12267. Epub 2013 Aug 14. Plant J. 2013. PMID: 23786378 Free PMC article.
-
Potential insect threats to pennycress, Thlaspi arvense (Brassicales: Brassicaceae), an emerging oilseed cover crop.J Insect Sci. 2024 Jul 1;24(4):17. doi: 10.1093/jisesa/ieae086. J Insect Sci. 2024. PMID: 39189128 Free PMC article. Review.
-
A draft genome of field pennycress (Thlaspi arvense) provides tools for the domestication of a new winter biofuel crop.DNA Res. 2015 Apr;22(2):121-31. doi: 10.1093/dnares/dsu045. Epub 2015 Jan 27. DNA Res. 2015. PMID: 25632110 Free PMC article.
Cited by
-
Root and Shoot Response to Nickel in Hyperaccumulator and Non-Hyperaccumulator Species.Plants (Basel). 2021 Mar 9;10(3):508. doi: 10.3390/plants10030508. Plants (Basel). 2021. PMID: 33803420 Free PMC article.
-
Genomic analysis of field pennycress (Thlaspi arvense) provides insights into mechanisms of adaptation to high elevation.BMC Biol. 2021 Jul 22;19(1):143. doi: 10.1186/s12915-021-01079-0. BMC Biol. 2021. PMID: 34294107 Free PMC article.
-
Natural variation and improved genome annotation of the emerging biofuel crop field pennycress (Thlaspi arvense).G3 (Bethesda). 2022 May 30;12(6):jkac084. doi: 10.1093/g3journal/jkac084. G3 (Bethesda). 2022. PMID: 35416986 Free PMC article.
-
Molecular tools enabling pennycress (Thlaspi arvense) as a model plant and oilseed cash cover crop.Plant Biotechnol J. 2019 Apr;17(4):776-788. doi: 10.1111/pbi.13014. Epub 2018 Oct 25. Plant Biotechnol J. 2019. PMID: 30230695 Free PMC article.
-
Two Arabidopsis promoters drive seed-coat specific gene expression in pennycress and camelina.Plant Methods. 2023 Dec 6;19(1):140. doi: 10.1186/s13007-023-01114-x. Plant Methods. 2023. PMID: 38053155 Free PMC article.
References
-
- Andersson, C. R. , Helliwell, C. A. , Bagnall, D. J. , Hughes, T. P. , Finnegan, E. J. , Peacock, W. J. , & Dennis, E. S. (2008). The FLX gene of Arabidopsis is required for FRI‐dependent activation of FLC expression. Plant and Cell Physiology, 49(2), 191–200. - PubMed
-
- Best, K. , & Mc Intyre, G. (1976). Studies on the flowering of Thlaspi arvense L. III. The influence of vernalization under natural and controlled conditions. Botanical Gazette, 137, 121–127. 10.1086/336850 - DOI
-
- Best, K. , & McIntyre, G. (1972). Studies on the flowering time of Thlaspi arvense L. I. The Influence of some environmental and genetic factors. Botanical Gazette, 133(4), 454–459. 10.1086/336670 - DOI
LinkOut - more resources
Full Text Sources
