

Positive and negative regulation of seed germination by the Arabidopsis GA hormone receptors, GID1a, b, and c
- PMID: 31245748
- PMCID: PMC6508844
- DOI: 10.1002/pld3.83
Positive and negative regulation of seed germination by the Arabidopsis GA hormone receptors, GID1a, b, and c
Abstract
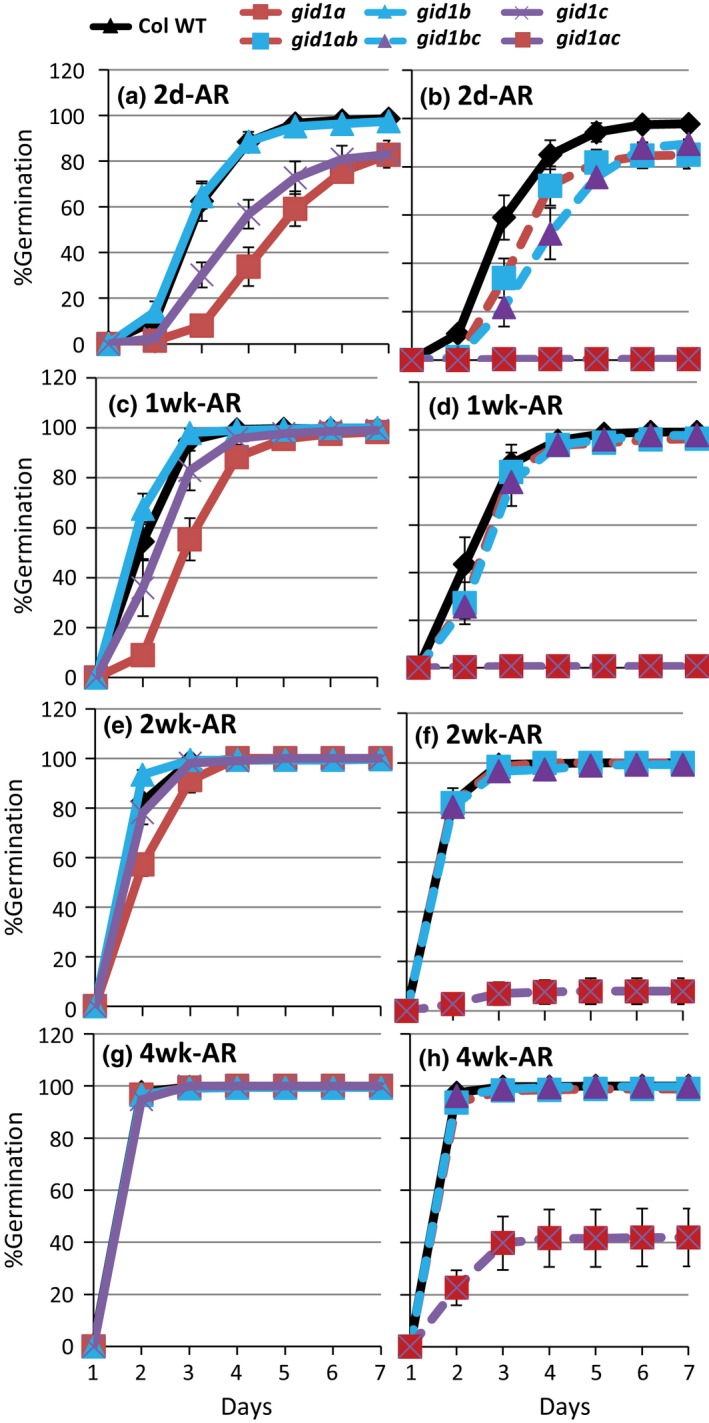
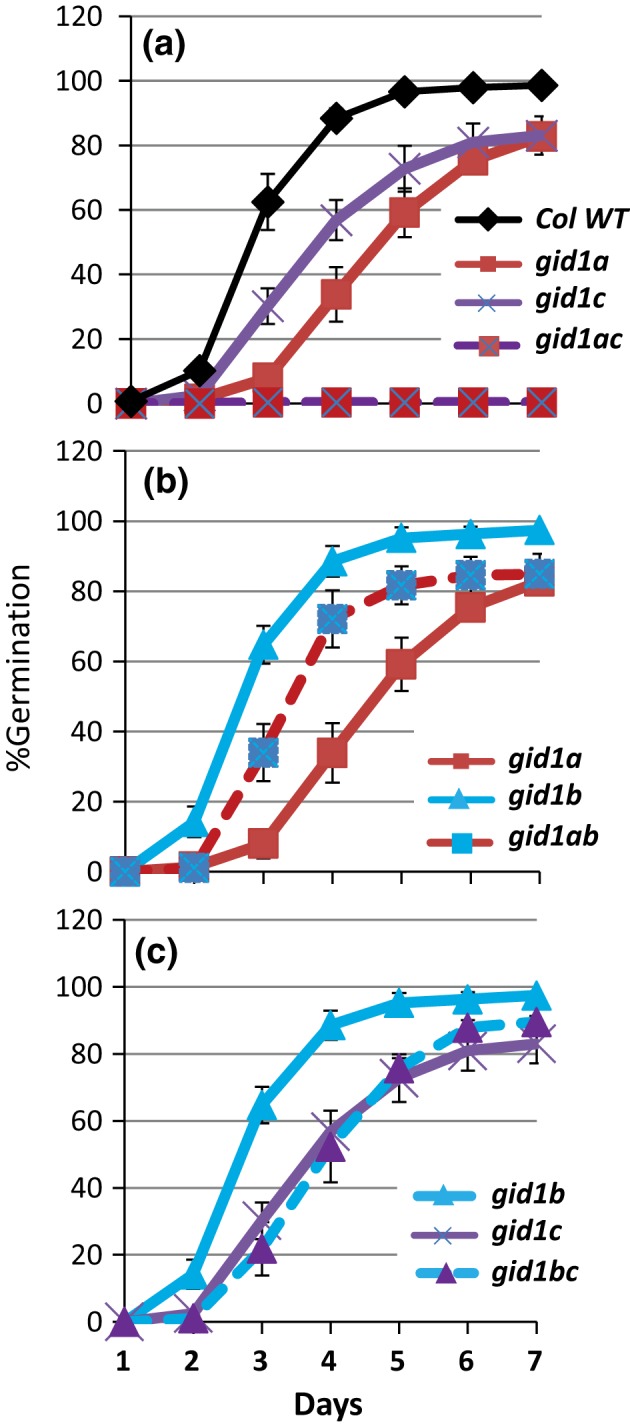
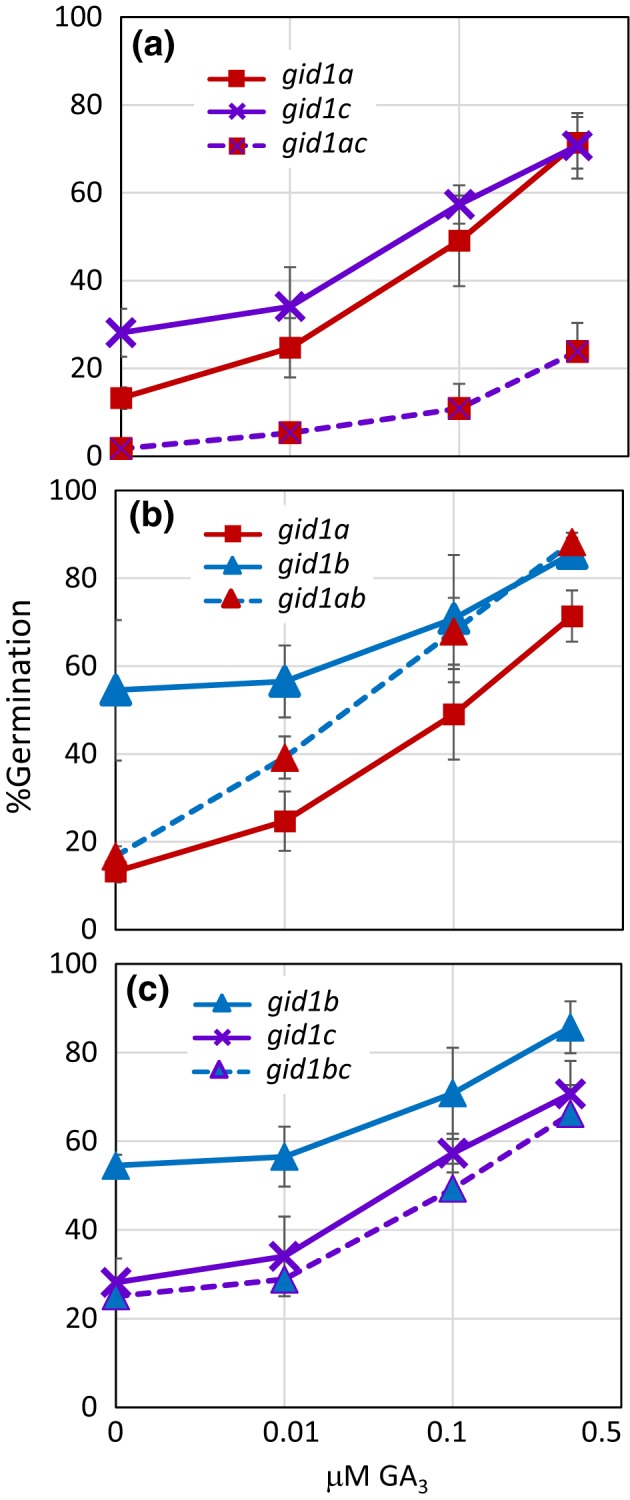
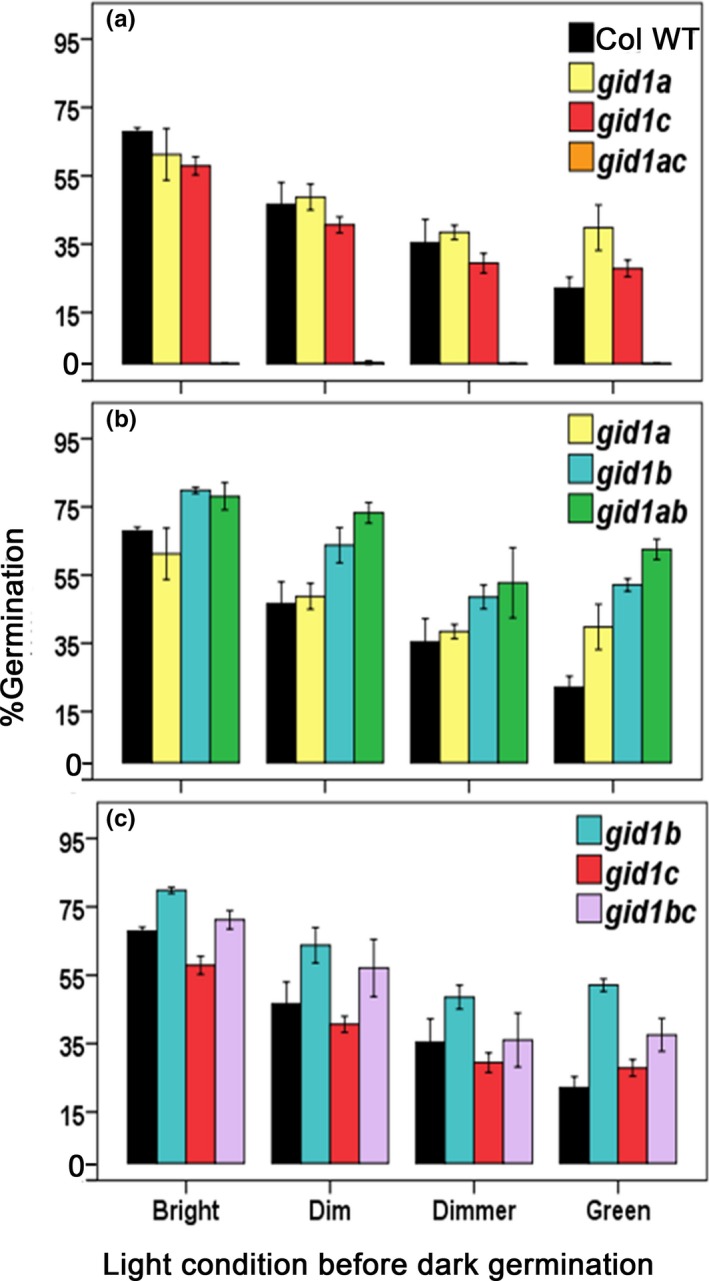
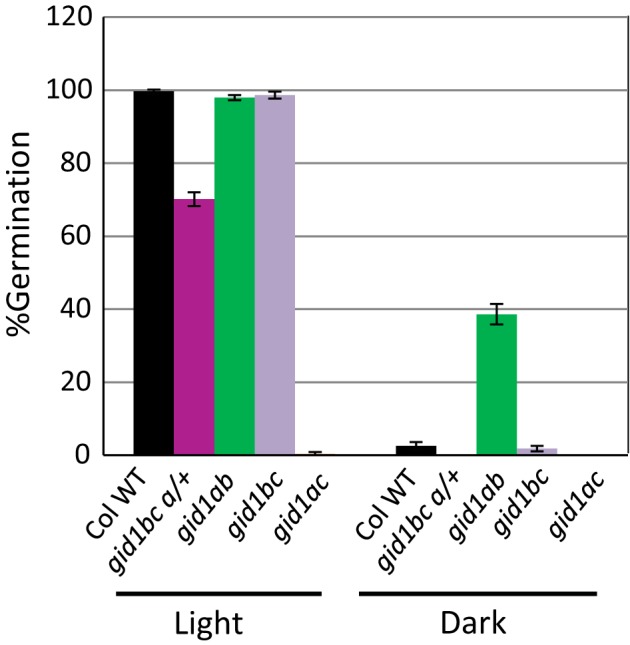
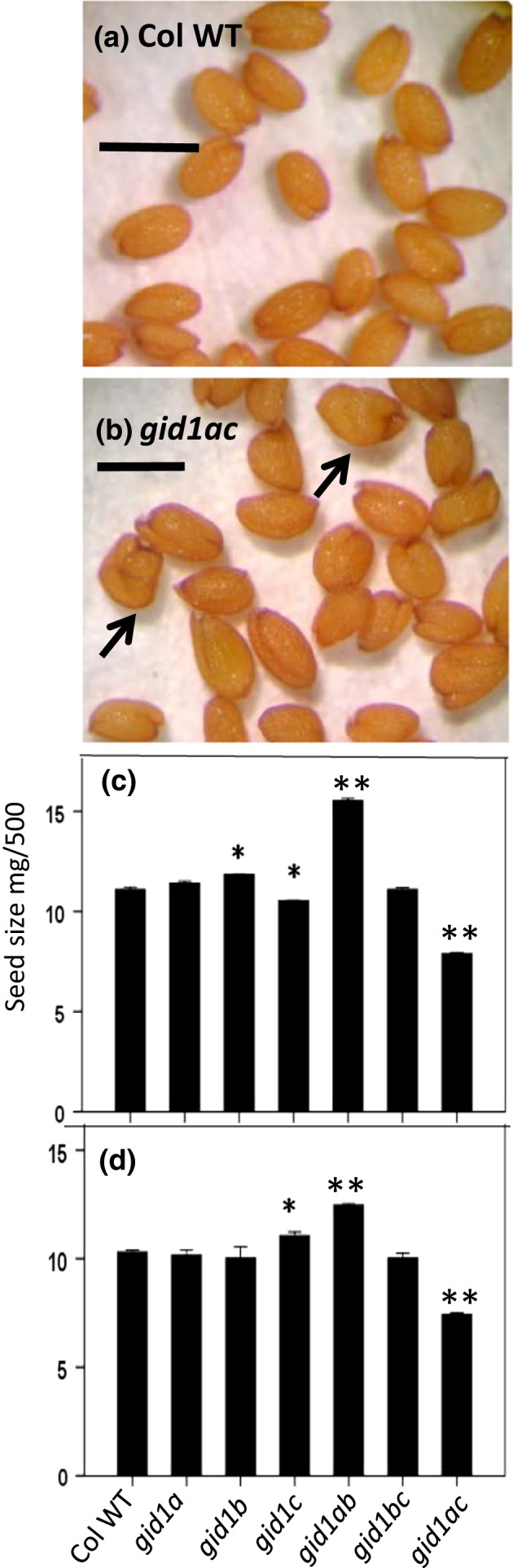
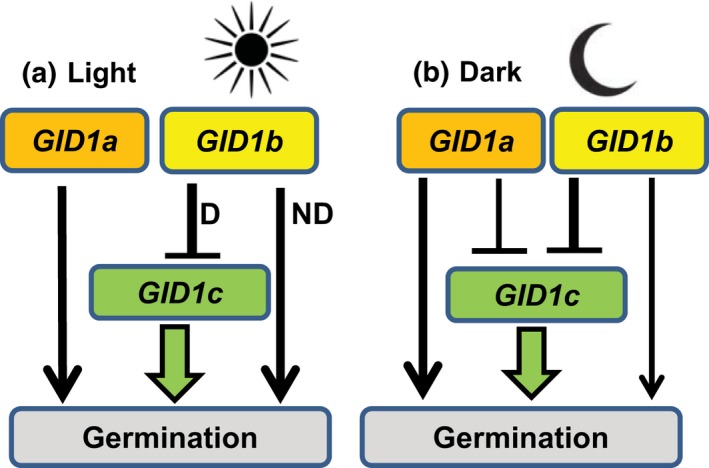
Epistasis analysis of gid1 single and double mutants revealed that GID1c is a key positive regulator of seed germination, whereas the GID1b receptor can negatively regulate germination in dormant seeds and in the dark. The GID1 GA receptors were expected to positively regulate germination because the plant hormone gibberellin (GA) is required for seed germination in Arabidopsis thaliana. The three GA hormone receptors, GID1a, GID1b, and GID1c, positively regulate GA responses via GA/GID1-stimulated destruction of DELLA (Asp-Glu-Leu-Leu-Ala) repressors of GA responses. The fact that the gid1abc triple mutant but not gid1 double mutants fail to germinate indicates that all three GA receptors can positively regulate non-dormant seed germination in the light. It was known that the gid1abc triple mutant fails to lose dormancy through the dormancy breaking treatments of cold stratification (moist chilling of seeds) and dry after-ripening (a period of dry storage). Previous work suggested that there may be some specialization of GID1 gene function during germination because GID1b mRNA expression was more highly induced by after-ripening, whereas GID1a and GID1c mRNA levels were more highly induced by cold stratification. In light-germinated dormant seeds, the gid1b mutation can partly rescue the germination efficiency of gid1a but not of gid1c seeds. Thus, GID1b can function as an upstream negative regulator GID1c, a positive regulator of dormant seed germination. Further experiments showed that GID1b can negatively regulate dark germination. Wild-type Arabidopsis seeds do not germinate well in the dark. The gid1b and gid1ab double mutants germinated much more efficiently than wild type, gid1c, or gid1ac mutants in the dark. The observation that the gid1ab double mutant also shows increased dark germination suggests that GID1b, and to some extent GID1a, can act as upstream negative regulators of GID1c. Since the gid1abc triple mutant failed to germinate in the dark, it appears that GID1c is a key downstream positive regulator of dark germination. This genetic analysis indicates that the three GID1 receptors have partially specialized functions in GA signaling.
Keywords: GID1; epistasis; germination; gibberellin; seed dormancy.
Figures
References
-
- Bewley, J. D. , Bradford, K. J. , Hilhorst, H. W. M. , & Nonogaki, H . (2013). SEEDS: Physiology of development, germination and dormancy. New York, NY, Springer; 10.1007/978-1-4614-4693-4 - DOI
-
- Chaisurisri, K. , Edwards, D. , & El‐Kassaby, Y. (1992). Genetic control of seed size and germination in Sitka spruce. Silvae Genet, 41, 348–355.
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
