

ARF5/MONOPTEROS directly regulates miR390 expression in the Arabidopsis thaliana primary root meristem
- PMID: 31245759
- PMCID: PMC6508847
- DOI: 10.1002/pld3.116
ARF5/MONOPTEROS directly regulates miR390 expression in the Arabidopsis thaliana primary root meristem
Abstract
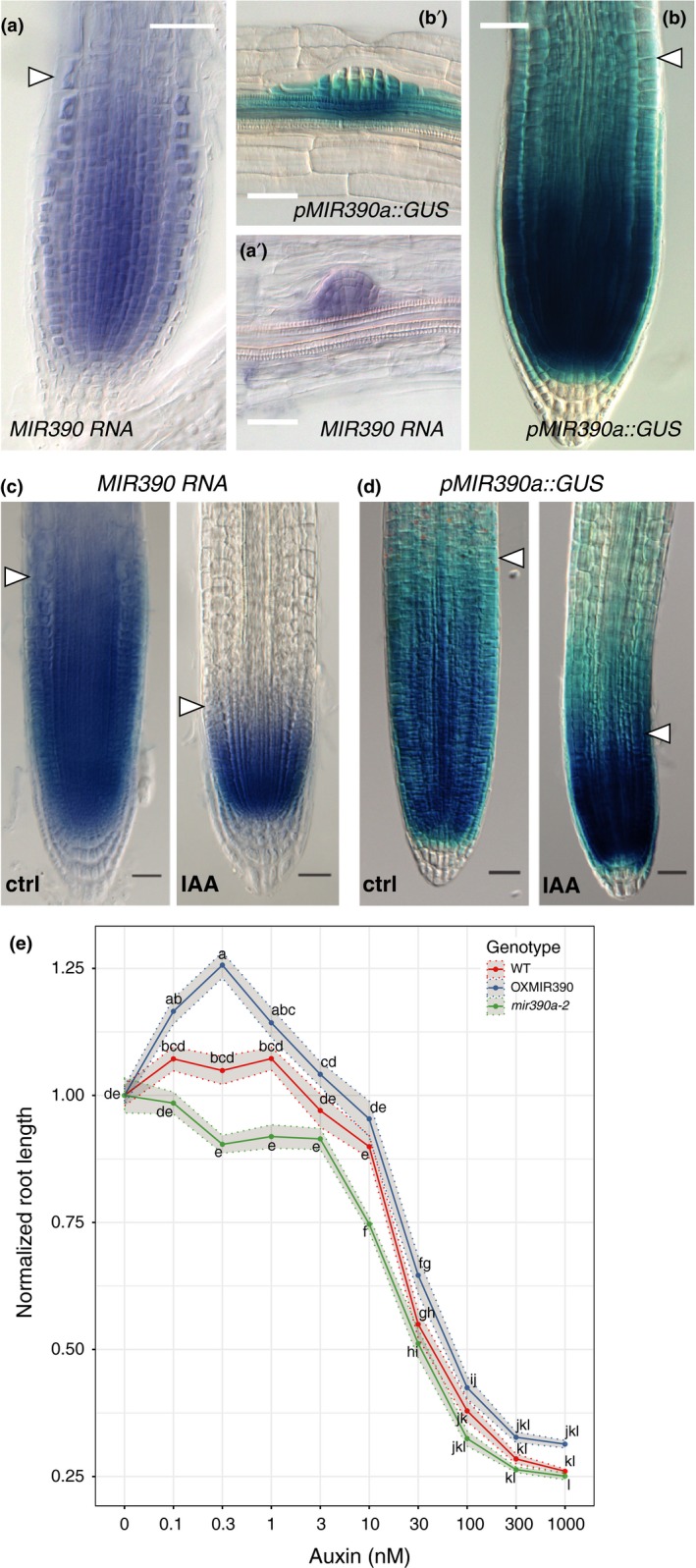
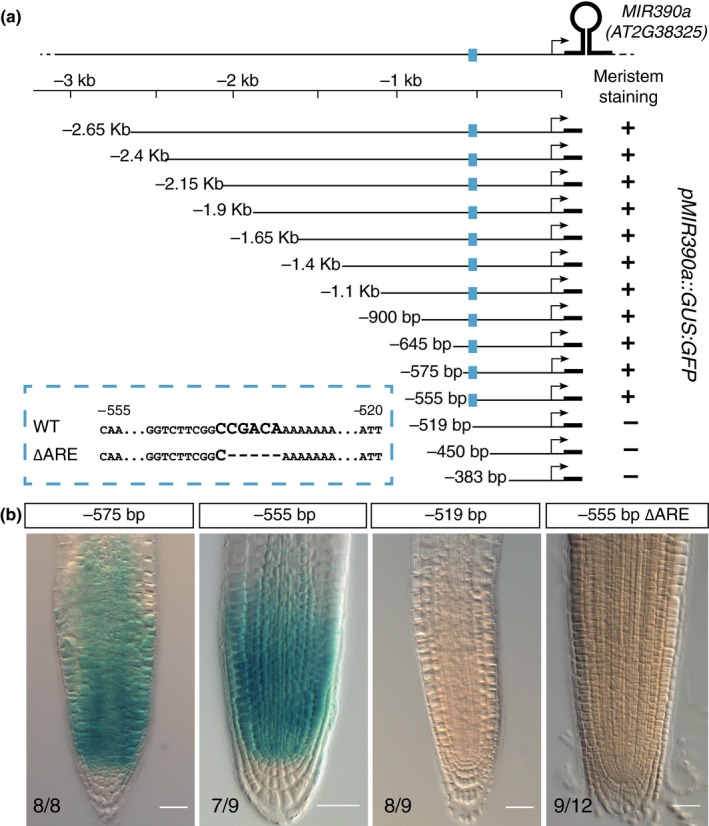
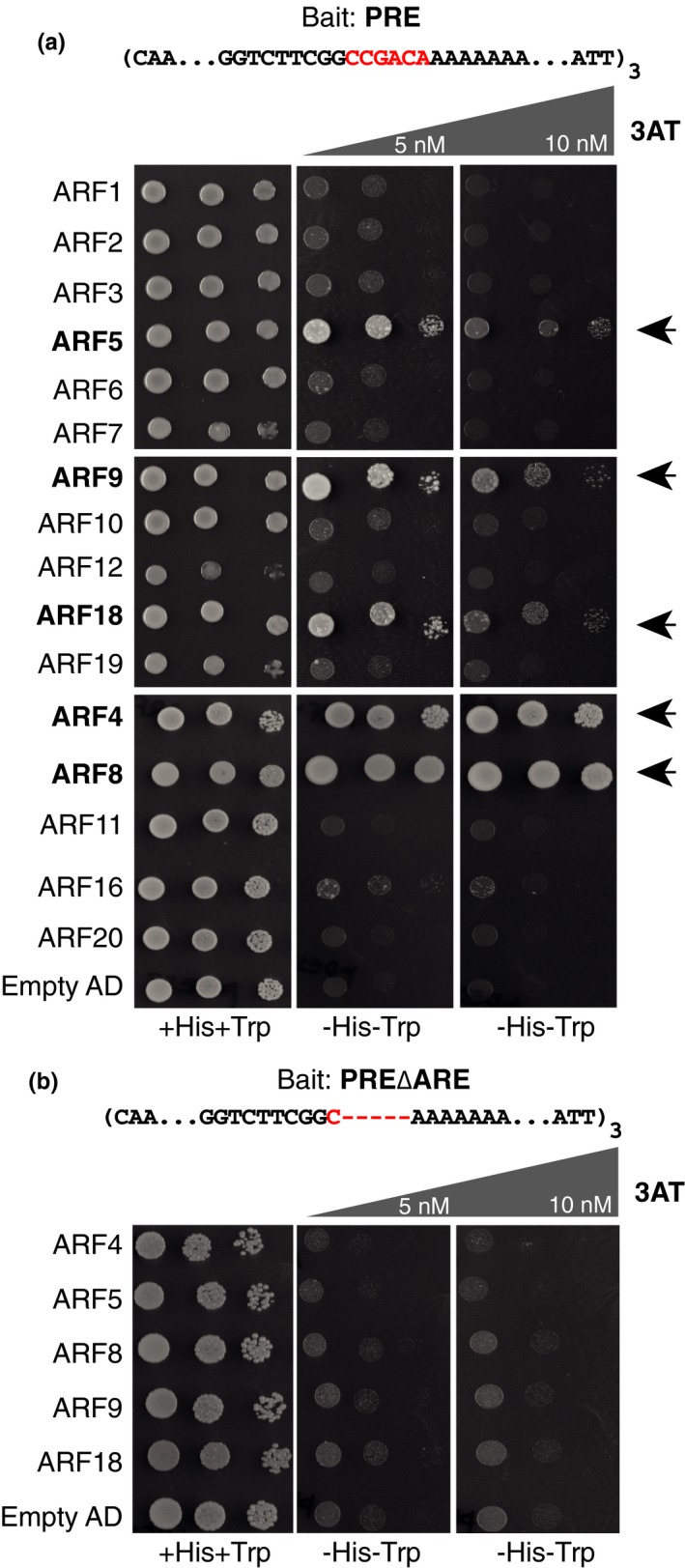
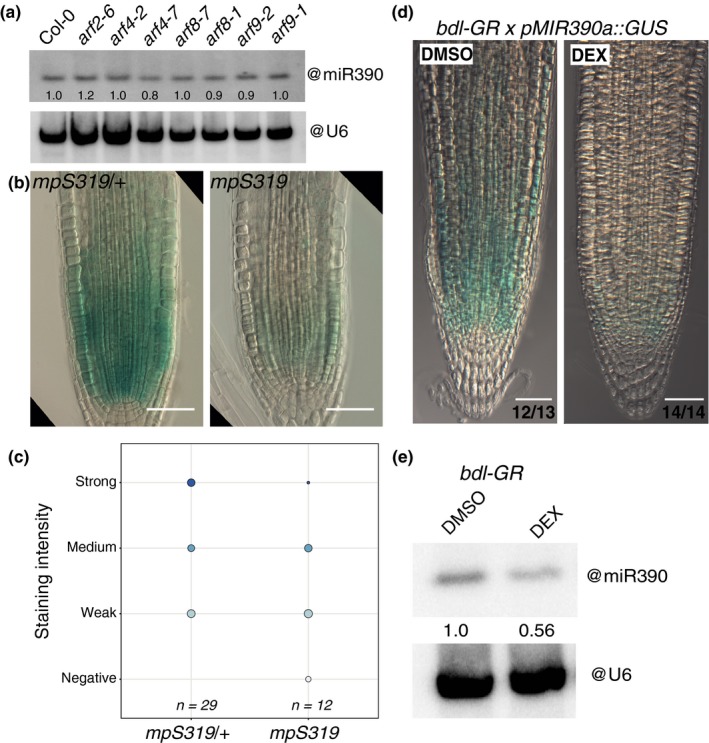
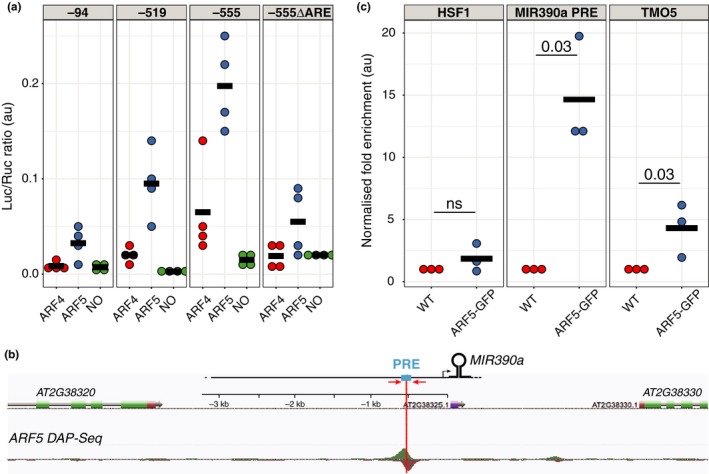
The root meristem is organized around a quiescent center (QC) surrounded by stem cells that generate all cell types of the root. In the transit-amplifying compartment, progeny of stem cells further divides prior to differentiation. Auxin controls the size of this transit-amplifying compartment via auxin response factors (ARFs) that interact with auxin response elements (AuxREs) in the promoter of their targets. The microRNA miR390 regulates abundance of ARF2, ARF3, and ARF4 by triggering the production of trans-acting (ta)-siRNA from the TAS3 precursor. This miR390/TAS3/ARF regulatory module confers sensitivity and robustness to auxin responses in diverse developmental contexts and organisms. Here, we show that miR390 is expressed in the transit-amplifying compartment of the root meristem where it modulates response to exogenous auxin. We show that a single AuxRE located in miR390 promoter is necessary for miR390 expression in this compartment and identify that ARF5/MONOPTEROS (MP) binds miR390 promoter via the AuxRE. We show that interfering with ARF5/MP-dependent auxin signaling attenuates miR390 expression in the transit-amplifying compartment of the root meristem. Our results show that ARF5/MP regulates directly the expression of miR390 in the root meristem. We propose that ARF5, miR390, and the ta-siRNAs-regulated ARFs modulate the response of the transit-amplifying region of the meristem to exogenous auxin.
Keywords: Arabidopsis; auxin; meristem; miRNA; root.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Aida, M. , Vernoux, T. , Furutani, M. , Traas, J. , & Tasaka, M. (2002). Roles of PIN‐FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo. Development, 129, 3965–3974. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
