

Structures of vertebrate Patched and Smoothened reveal intimate links between cholesterol and Hedgehog signalling
- PMID: 31247512
- PMCID: PMC6744280
- DOI: 10.1016/j.sbi.2019.05.015
Structures of vertebrate Patched and Smoothened reveal intimate links between cholesterol and Hedgehog signalling
Abstract
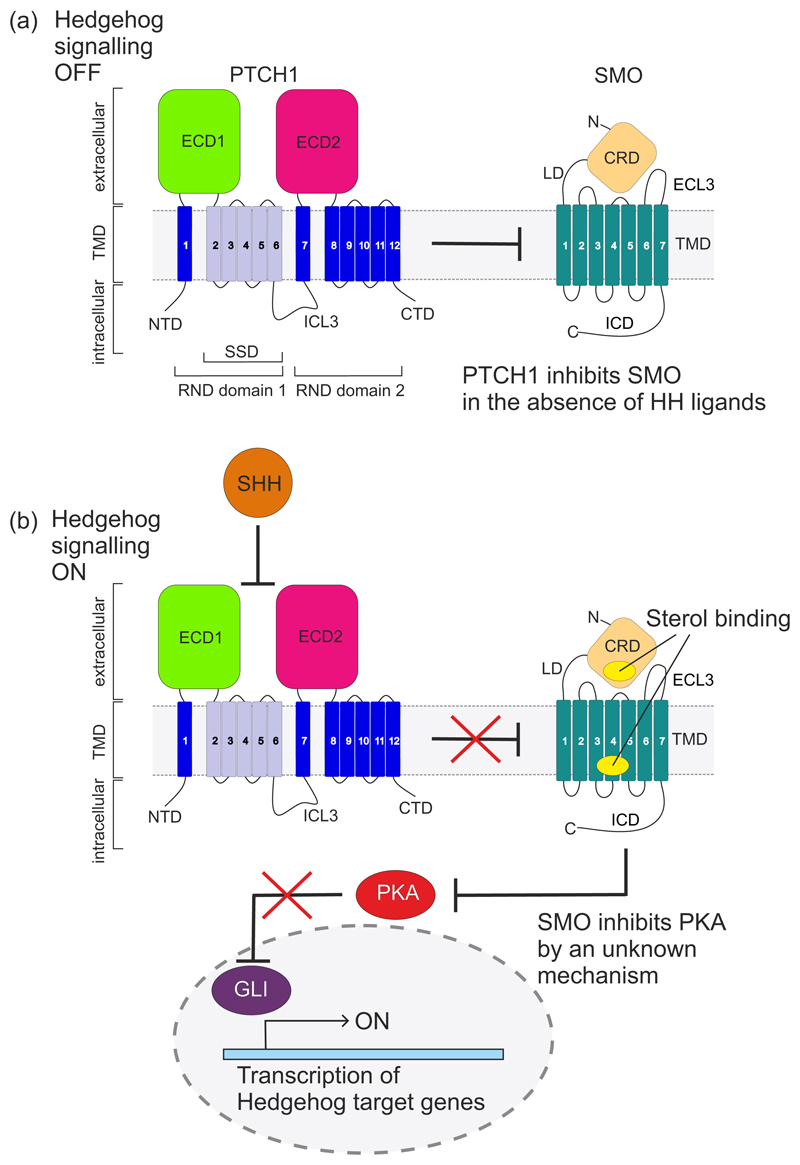
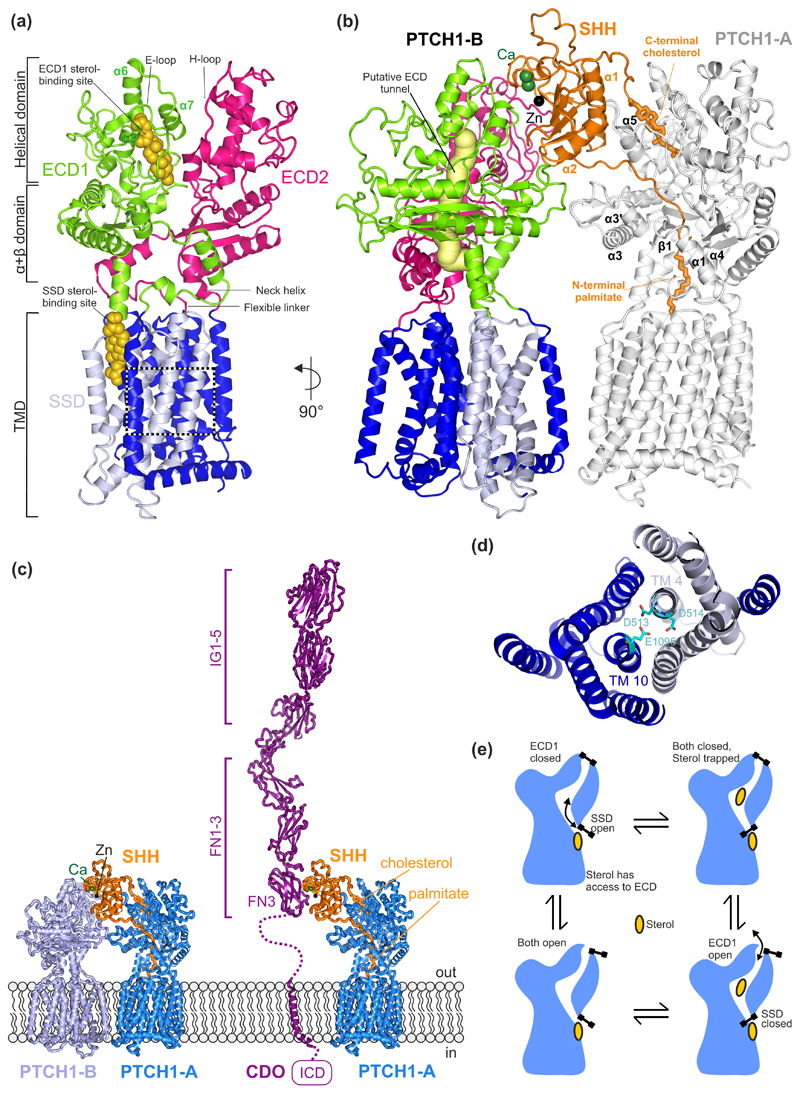
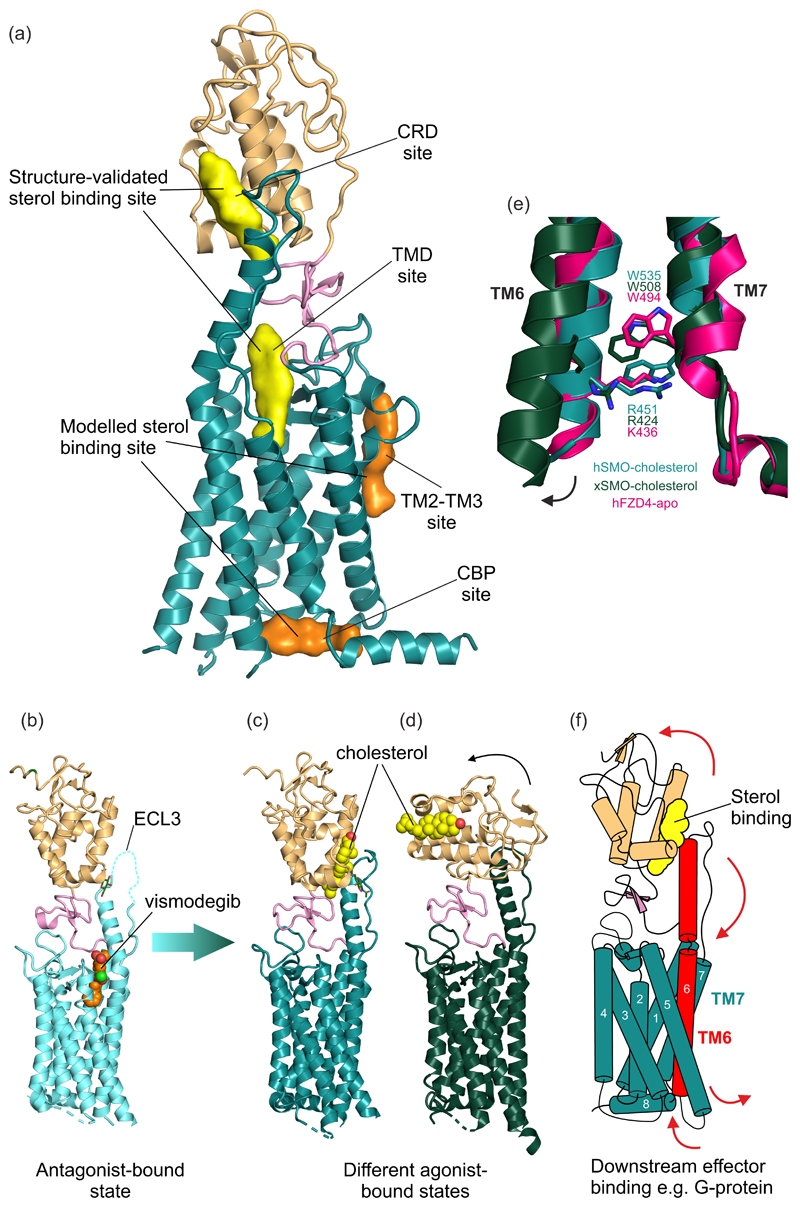
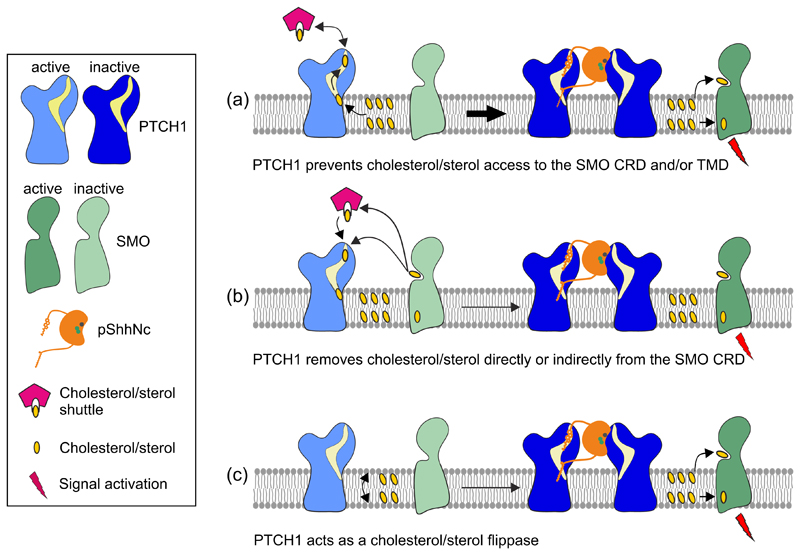
The Hedgehog (HH) signalling pathway is a cell-cell communication system that controls the patterning of multiple tissues during embryogenesis in metazoans. In adults, HH signals regulate tissue stem cells and regenerative responses. Abnormal signalling can cause birth defects and cancer. The HH signal is received on target cells by Patched (PTCH1), the receptor for HH ligands, and then transmitted across the plasma membrane by Smoothened (SMO). Recent structural and biochemical studies have pointed to a sterol lipid, likely cholesterol itself, as the elusive second messenger that communicates the HH signal between PTCH1 and SMO, thus linking ligand reception to transmembrane signalling.
Crown Copyright © 2019. Published by Elsevier Ltd. All rights reserved.
Figures
References
-
- Lee RT, Zhao Z, Ingham PW. Hedgehog signalling. Development. 2016;143:367–372. - PubMed
-
- Pepinsky RB, Zeng C, Wen D, Rayhorn P, Baker DP, Williams KP, Bixler SA, Ambrose CM, Garber EA, Miatkowski K, et al. Identification of a palmitic acid-modified form of human Sonic hedgehog. J Biol Chem. 1998;273:14037–14045. - PubMed
-
- Porter JA, Ekker SC, Park WJ, von Kessler DP, Young KE, Chen CH, Ma Y, Woods AS, Cotter RJ, Koonin EV, et al. Hedgehog patterning activity: role of a lipophilic modification mediated by the carboxy-terminal autoprocessing domain. Cell. 1996;86:21–34. - PubMed
-
- Porter JA, Young KE, Beachy PA. Cholesterol modification of hedgehog signaling proteins in animal development. Science. 1996;274:255–259. - PubMed
-
- Marigo V, Davey RA, Zuo Y, Cunningham JM, Tabin CJ. Biochemical evidence that patched is the Hedgehog receptor. Nature. 1996;384:176–179. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- C20724/A16135/CRUK_/Cancer Research UK/United Kingdom
- R01 GM112988/GM/NIGMS NIH HHS/United States
- R35 GM118082/GM/NIGMS NIH HHS/United States
- 203726/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- C20724/A26752/CRUK_/Cancer Research UK/United Kingdom
- R01 GM106078/GM/NIGMS NIH HHS/United States
- 26752/CRUK_/Cancer Research UK/United Kingdom
- 14414/CRUK_/Cancer Research UK/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- 203852/Z/16/2/WT_/Wellcome Trust/United Kingdom
- C20724/A14414/CRUK_/Cancer Research UK/United Kingdom
- 647278/ERC_/European Research Council/International
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
